Метаболизм жиров
Содержание:
Клиническое значение
При дефектах в гене фитаноил-СоА-гидроксилазы развивается синдром Рефсума. Причиной этого заболевания является невозможность α-окисления фитановой кислоты. У человека в норме эта кислота образуется из полипренильного растительного спирта фитола, который попадает в организм в эфирной форме вместе с хлорофиллом, содержащимся в пище. Хотя лишь малая доля фитола, поступающего с пищей, окисляется до фитановой кислоты, она дополнительно поступает в организм вместе с некоторыми животными жирами и молочными продуктами. Поскольку α-окисление при дефектном ферменте невозможно, фитановая кислота подвергается β-окислению с образованием трёх молекул , трёх молекул ацетил-СоА и одной молекулы . Эта болезнь, описанная Рефсумом в 1946 году, сопровождается серьёзными повреждениями нервов и мозга, накоплением липидов и ранней смертью.
Регуляция
Скорость регуляции процесса β-окисления включает несколько факторов:
- Соотношений АТФ/АМФ и НАДH/НАД+, так же, как и скорость реакций ЭТЦ и общего пути катаболизма;
- состояния голодания или сытости (то есть соотношения инсулин — глюкагон);
- активности регуляторного фермента карнитин-пальмитоилтрансферазы I (CPTI);
- доступности субстрата — жирных кислот;
- потребности клетки в энергии;
- доступности кислорода.
Скорость β-окисления зависит также от активности фермента карнитин-пальмитоилтрансферазы I (CPTI). В печени этот фермент ингибируется малонил-КоА, веществом, образующимся при биосинтезе жирных кислот.
В мышцах карнитин-пальмитоилтрансфераза I (CPTI) также ингибируется малонил-КоА. Хотя мышечная ткань не синтезирует жирные кислоты, в ней имеется изофермент ацетил-КоА-карбоксилазы, синтезирующий малонил-КоА для регуляции β-окисления. Данный изофермент фосфорилируется протеинкиназой А, которая активируется в клетках под действием адреналина, и АМФ-зависимой протеинкиназой и таким образом происходит его ингибирование; концентрация малонил-КоА снижается. Вследствие этого, при физической работе, когда в клетке появляется АМФ, под действием адреналина активируется β-окисление, однако, его скорость зависит ещё и от доступности кислорода. Поэтому β-окисление становится источником энергии для мышц только через 10-20 минут после начала физической нагрузки (так называемые аэробные нагрузки), когда приток кислорода к тканям увеличивается.
Примечания
- ↑ Строев Е. А. Биологическая химия: Учебник для фармац. ин-тов и фармац. фак. мед. ин-тов. — М.: Высшая школа, 1986. — 479 с.
- ↑ Е.С. Северин. Биохимия. — М: ГЭОТАР-МЕД, 2004. — 779 с. — ISBN 5-9231-0254-4.
- ↑ Березов Т. Т., Коровкин Б. Ф. Биологическая химия. — М.: Медицина, 1998. — 704 с. — ISBN 5-225-02709-1.
- ↑ , p. 943.
- Knoop, Franz (1904). «Der Abbau aromatischer Fettsäuren im Tierkörper». Beitr Chem Physiol Pathol 6: 150–162. Проверено 2 March 2015.
- Voet, Donald. Fundamentals of Biochemistry Life at the Molecular Level / Donald Voet, Voet, Pratt. — New York City : John Wiley & Sons, Inc., 2013. — P. 582–584. — ISBN 1118129180.
- ↑ Р.Марри, Д.Греннер, П. Мейес, В. Родуэлл. Биохимия человека. — М.: Мир, 1993. — Т. I. — 384 с. — ISBN 5-03-001774-7.
- ↑ Нельсон Д., Кокс М. Основы биохимии Ленинджера. — М.: БИНОМ, 2011. — Т. II.
- Кольман. Я., Рём К. Г. Наглядная биохимия. — М.: Мир, 2011. — 469 с. — ISBN 5-03-003304-1.
- Биологическая химия с упражнениями и задачами / Под ред. С.Е. Северина. — М.: ГЭОТАР-Медиа, 2011. — 624 p. — ISBN 9785970417553.
- P. Bowen, C. S. N. Lee, H. U. Zellweger, R. Lindenburg. A familial syndrome of multiple congenital defects. Bulletin of the Johns Hopkins Hospital, 1964; 114: 402.
- OMIM
Силовые тренировки
Задача кардио запустить окислительные процессы, а если после
силовой тренировки у нас истощены запасы креатин фосфата, гликогена — это будет являться
отличной почвой для окисления жиров липолиза.
Есть два термина обозначающих накопление жиров и расщепление
— липогенез и липолиз соответственно. Если говорить в двух словах об отличии расщепления
и накопления жиров, то липолиз вовсе не гарантирует нам снижение жирового
компонента, а окисление жиров гарантирует.
Жир – это самая энергоемкая единица в нашем организме. В 1
гр жира содержится 9 ккал, тогда как в белках и углеводах только по 4 гр. Тем не менее, организм сложней заставить использовать жиры в качестве энергии, потому что они
отложены у нас в жировом депо, жировых клетках.
Мы знаем, что организм пытается на всем сэкономить, поэтому
пока нет нужды извлекать эти жиры он этого делать и не будет.

Как похудеть в таком случае?
Все очень просто. Необходимо тратить энергии больше чем мы
ее потребляем и на наш взгляд это можно сделать несколькими способами.
- »Ничего не меняя снизить количество потребляемых
калорий - »Повысить уровень своей двигательной активности,
записавшись в спортивную секцию или тренажерный зал - »Меньше есть и больше двигаться – самый
эффективный способ
Нормализованное питание плюс тренировки ровняется
ускоренному процессу жиросжигания. Еще как бонус вы получите избавление от
мышечной атрофированности и ряд приятных моментов присущих нормальной
двигательной активности. Ничего сверхчеловеческого делать мы не будем, для
снижения жирового компонента это и не нужно.
Итак, мы нормализовали питание, начали заниматься по
адекватной схеме тренировок и после силовой тренировки добавили, к примеру,
получасовое кардио либо ходьбу в быстром темпе. Что же происходит в это время в
нашем организме?
Стресс-реакцию при силовых тренировках. Эта
реакция мобилизирует наши энергетические ресурсы под воздействием
катаболических гормонов и запускает процесс адаптации. К слову всем знакомая
фраза «делай базу» несет на себе именно смысл в том, что бы достичь
стресс-реакции. Для здорового человека необходимы упражнения глобального характера,
которые нагрузят весь наш организм, запустят процесс стресс-реакции, адаптации.
Особенности окисления в пероксисомах
Жирные кислоты представляют собой углеродные цепи различной длины (от 4 до 36 атомов), которых по химической природе относят к карбоновым кислотам. Эти цепи могут быть как разветвленными, так и не разветвленными и содержать разное количество двойных связей. Если последние полностью отсутствуют, жирные кислоты называют насыщенными (характерно для многих липидов животного происхождения), а в противном случае – ненасыщенными. По расположению двойных связей жирные кислоты подразделяют на мононенасыщенные и полиненасыщенные.
Большинство цепей содержит четное число атомов углерода, что связано с особенностью их синтеза. Однако есть соединения с нечетным количеством звеньев. Окисление этих двух типов соединений несколько отличается.
https://www.youtube.com/watch?v=userlifehackertv
К самому простому типу энергетической утилизации жирных кислот относят β-окисление не имеющих двойных связей цепей, в которых количество углеродных звеньев четное. В качестве субстрата для этого процесса, как уже выше отмечалось, выступает ацил коэнзима А.
Процесс β-окисления жирных кислот состоит из 4 реакций:
- Дегидрирование – отщепление водорода от β-углеродного атома с возникновением двойной связи между звеньями цепи, находящимися в α и β-положениях (первый и второй атомы). В результате образуется еноил-CoA. Ферментом реакции является ацил-CoA-дегидрогеназа, которая действует в комплексе с кофермента ФАД (последний восстанавливается до ФАДН2).
- Гидратация – присоединение молекулы воды к еноил-CoA, в результате чего образуется L-β-гидроксиацил-CoA. Осуществляется еноил-CoA-гидратазой.
- Дегидрирование – окисление продукта предыдущей реакции НАД-зависимой дегидрогеназой с образованием β-кетоацил-коэнзима А. При этом происходит восстановление НАД до НАДН.
- Расщепление β-кетоацил-CoA до ацетил-CoA и укороченного на 2 атома углерода ацил-CoA. Реакция осуществляется под действием тиолазы. Обязательным условием является присутствие свободного HS-CoA.
Цикличное повторение всех стадий осуществляется до тех пор, пока вся углеродная цепочка жирной кислоты не превратится в молекулы ацетил-коэнзима А.
Окисление ненасыщенных жирных кислот имеет некоторые особенности. Так, сложность окисления цепей с двойными связями заключается в том, что последние не могут подвергаться воздействию еноил-CoA-гидратазы из-за того, что находятся в цис-положении. Эта проблема устраняется еноил-CoA-изомеразой, благодаря которой связь приобретает транс-конфигурацию.
Иногда для продолжения процесса недостаточно еноил-CoA-изомеразы. Это касается цепей, в которых присутствует конфигурация цис9-цис12 (двойные связи при 9-м и 12-м атомах углерода). Здесь помехой является не только конфигурация, но и положение двойных связей в цепи. Последнее исправляется ферментом 2,4-диеноил-CoA-редуктазой.
Второе важное различие заключается в том, что некоторые ферменты пероксисом специфичны к определенным малораспространенным жирным кислотам и отсутствуют в митохондриальном матриксе. Особенность пероксисом клеток печени заключается в том, что там отсутствует ферментный аппарат цикла Кребса
Поэтому в результате бета-окисления образуются короткоцепочечные продукты, которые для окисления транспортируются в митохондрии
Особенность пероксисом клеток печени заключается в том, что там отсутствует ферментный аппарат цикла Кребса. Поэтому в результате бета-окисления образуются короткоцепочечные продукты, которые для окисления транспортируются в митохондрии.
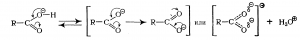
Химические свойства
Химические свойства Ж. к. определяются свойствами их COOH-групп и углеводородного радикала. В COOH-группе связь O—H ослаблена за счет смещения электронной плотности в двойной C=O связи к кислороду, и поэтому протон может быть легко отщеплен. Это приводит к появлению стабильного аниона к-ты:
Сродство карбонилового остатка к электронам может быть частично удовлетворено за счет соседней метиленовой группы, водородные атомы к-рой наиболее активны по сравнению с остальными. Константа диссоциации COOH-группы Ж. к. равна 10-4—10-5 М, т. е. ее величина гораздо ниже, чем у неорганических к-т. Наиболее сильной из Ж. к. является муравьиная к-та. COOH-группа Ж. к. обладает способностью реагировать в водных р-рах с щелочноземельными металлами. Соли высших Ж. к. с этими металлами называются мылами (см.). Мыла обладают свойствами поверхностно-активных веществ — детергентов (см.). Натриевые мыла твердые, калиевые — жидкие. Гидроксил COOH-групп Ж. к. может быть легко замещен на галоген с образованием галогенангидридов, которые широко используются в органических синтезах. При замещении галогена остатком другой к-ты образуются ангидриды Ж. к., при замещении остатком спирта — их сложные эфиры, аммиаком — амиды, гидразином — гидразиды. Наиболее распространены в природе сложные эфиры трехосновного спирта глицерина и высших Ж. к. — жиры (см.). Водород альфа-углеродного атома Ж. к. может быть легко замещен галогеном с образованием галогенсодержащих Ж. к. Непредельные Ж. к. могут существовать в виде цис- и транс-изомеров. Большинство природных ненасыщенных Ж. к. имеют цис-конфигурацию (см. Изомерия). Степень ненасыщенности Ж. к. определяют йодометрическим титрованием двойных связей. Процесс превращения ненасыщенных Ж. к. в насыщенные получил название гидрогенизации, обратный процесс— дегидрогенизации (см. Гидрогенизация).
Природные Ж. к. получают путем гидролиза жиров (их омыления) с последующей дробной перегонкой или хроматографическим разделением освободившихся Ж. к. Неприродные Ж. к. получают путем окисления углеводородов; реакция протекает через стадию образования гидроперекисей и кетонов.
Энергетический баланс процесса
В результате переноса электронов по ЭТЦ от ФАДH2 и НАДH синтезируется по 5 молекул АТФ (2 от ФАДH2, и 3 от НАДH). В случае окисления пальмитиновой кислоты проходит 7 циклов β-окисления (16/2-1=7), что ведёт к образованию 5•7=35 молекул АТФ. В процессе β-окисления пальмитиновой кислоты образуется n молекул ацетил-КоА, каждая из которых, при полном сгорании в цикле трикарбоновых кислот, даёт 12 молекул АТФ, а 8 молекул дадут 12•8 = 96 молекул АТФ.
Таким образом, всего при полном окислении пальмитиновой кислоты образуется 35+96=131 молекула АТФ. Однако с учётом одной молекулы АТФ, которая гидролизуется до АМФ, то есть тратятся 2 макроэргические связи или две АТФ, в самом начале на процесс активирования (образования пальмитоил-CоА) общий энергетический выход при полном окислении одной молекулы пальмитиновой кислоты в условиях животного организма составит 131-2=129 молекул.
Суммарное уравнение окисления пальмитиновой кислоты выглядит следующим образом:
- C15H31CO−SCoA+7FAD++7NAD++7H2O+7HS−CoA→8CH3CO−SCoA+7FADH2+7NADH{\displaystyle {\mathsf {C_{15}H_{31}CO-SCoA+7FAD^{+}+7NAD^{+}+7H_{2}O+7HS-CoA\rightarrow 8CH_{3}CO-SCoA+7FADH_{2}+7NADH}}}
Формула для расчёта общего количества АТФ которые генерируются в результате процесса β-окисления:
- (n2⋅12)+((n2−1)⋅5){\displaystyle {\left}}
где n — количество атомов углерода в молекуле жирной кислоты.
Энергетический расчёт β-окисления для некоторых жирных кислот представлен в виде таблицы.
| Жирная кислота | Кол-во молекул АТФ генерируемых на 1 молекулу жирной кислоты | Кол-во затраченных молекул АТФ | Общий энергетический выход молекул АТФ |
|---|---|---|---|
| Каприловая кислота C7H15COOH | 63 | 2 | 63-2=61 |
| Лауриновая кислота С11Н23COOH | 97 | 2 | 97-2=95 |
| Миристиновая кислота С13Н27СООН | 114 | 2 | 114-2=112 |
| Пентадециловая кислота С14Н29СООН | 122,5 | 2 | 122,5-2=120,5 |
| Пальмитиновая кислота С15Н31СООН | 131 | 2 | 131-2=129 |
| Маргариновая кислота С16Н33СООН | 139,5 | 2 | 139,5-2=137,5 |
| Стеариновая кислота С17Н35СООН | 148 | 2 | 148-2=146 |
| Арахиновая кислота С19Н39СООН | 165 | 2 | 165-2=163 |
Во многих тканях окисление жирных кислот — важный источник энергии. Это ткани с высокой активностью ферментов цикла Кребса и цепи переноса электронов — клетки красных скелетных мышц, сердечная мышца (миокард), почки. Например, эритроциты, в которых отсутствуют митохондрии, не могут окислять жирные кислоты. В то же время жирные кислоты не могут служить источником энергии для мозга и других нервных тканей, так как они не проходят через гематоэнцефалический барьер, вследствие их гидрофобных свойств. Скорость обмена жирных кислот в нервных тканях на порядок ниже чем, например, в скелетных мышцах. Поэтому в таких ситуациях, особенно при длительном голодании, печень перерабатывает около 50 % поступающих в неё жирных кислот в другие источники энергии — кетоновые тела, которые может утилизировать нервная ткань.
Химические свойства жиров
Химические свойства жиров обусловлены наличием:
- сложных эфирных связей;
- двойных связей в углеводородных радикалах жирных кислот;
- наличием глицерина в составе жира.
Гидролиз, или омыление
В зависимости от условий гидролиз бывает:
- кислотный (в присутствии кислоты в качестве катализатора);
- щелочной (под действием щелочей);
- водный (без катализатора, при высоких t и P);
- ферментативный (происходит в живых организмах).
1. Кислотный гидролиз
Жирам как сложным эфирам свойственна обратимая реакция гидролиза, катализируемая минеральными кислотами: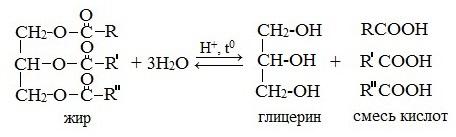
2. Щелочной гидролиз (реакция Шевреля)
При участии щелочей гидролиз жиров происходит необратимо — щелочи превращают образующиеся кислоты в соли и тем самым устраняют возможность взаимодействия кислот с глицерином. Продуктами в этом случае являются мыла — соли высших карбоновых кислот и щелочных металлов: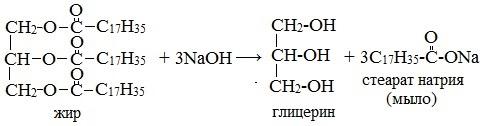
Натриевые соли — твердые мыла, калиевые — жидкие. Реакция щелочного гидролиза жиров, и вообще всех сложных эфиров, называется также омылением.
3. Водный гидролиз
Промышленный метод гидролиза жиров заключается в обработке их водяным паром при температуре 200С под давлением.
4. Ферментативный гидролиз
В организмах человека и животных жиры, поступающие в составе пищи, подвергаются гидролитическому расщеплению с участием специальных ферментов – липаз.
Гидрогенизация (гидрирование) жидких жиров
Жидкие жиры превращают в твердые путем реакции гидрогенизации (каталитического гидрирования). При этом водород присоединяется по двойной связи, содержащейся в углеводородном радикале молекул масел:
Продукт гидрогенизации масел — твердый жир (искусственное сало, саломас – сало из масла). Маргарин — пищевой жир, состоит из смеси гидрогенизированных масел (подсолнечного, кукурузного, хлопкого и др.), животных жиров, молока и вкусовых добавок (соли, сахара, витаминов и др.).
В условиях процесса гидрогенизации масел (высокая температура, металлический катализатор) происходит изомеризация части кислотных остатков, содержащих цис-связи С=С, в более устойчивые транс-изомеры. Повышенное содержание в маргарине (особенно, в дешевых сортах) остатков транс-ненасыщенных кислот увеличивает опасность атеросклероза, сердечно-сосудистых и других заболеваний.
Реакции окисления и полимеризации (для жидких ненасыщенных жиров)
Жиры, содержащие остатки ненасыщенных кислот (высыхающие масла), под действием кислорода воздуха окисляются и полимеризуются.
При длительном хранении жиры портятся (прогоркают). Под действием воздуха, света и микроорганизмов происходит частичный гидролиз жиров с образованием свободных жирных кислот и продуктов их превращения, обычно имеющих неприятный запах и вкус. Срок годности жиров увеличивается при низкой температуре и в присутствии консервантов (чаще всего поваренной соли).
Рубрики: Жиры Теги: Жиры
БИОХИМИЯ — Основные питательные вещества человека — 2016 год
2.4.2. Катаболизм жиров
Использование организмом жира в качестве источника энергии начинается с его выхода из жировых депо в кровяное русло. Этот процесс называется мобилизацией жира. Мобилизация жира ускоряется под влиянием симпатической нервной системы и гормона адреналин.
Основные превращения жира происходят в печени, где имеются активные ферменты жирового обмена.
В печени жир прежде всего подвергается гидролизу и превращается, так же, как и в кишечнике, в глицерин и жирные кислоты.
Образовавшийся глицерин легко переходит в фосфоглпцерпновый альдегид, который является также промежуточным продуктом распада углеводов и поэтому вовлекается в углеводный обмен.
Жирные кислоты вначале активируются с использованием энергии АТФ и связываются со своим переносчиком — коферментом А (комплекс «жирная кислота — кофермент А» называется ацилкоферментом А):
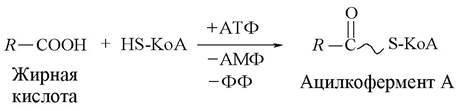
Затем ацилкофермент А с помощью еще одного переносчика — карнитина поступает в митохондрии, где происходит окисление жирной кислоты.
Окисление жирных кислот протекает в два этапа.
На первом этапе, называемом β — окислением (это название обусловлено тем, что окислению подвергается атом С жирной кислоты, находящийся в р!!!-положении углеродной цепи), от жирной кислоты, связанной с коферментом А, дважды отщепляются по два атома водорода, которые затем по дыхательной цепи передаются на молекулярный кислород. В итоге образуется вода, и за счет выделяющейся при этом энергии осуществляется синтез пяти молекул АТФ. Завершается р-окисление отщеплением от жирной кислоты двухуглеродного фрагмента в виде ацетилкофермента А (ацетил — КоА):
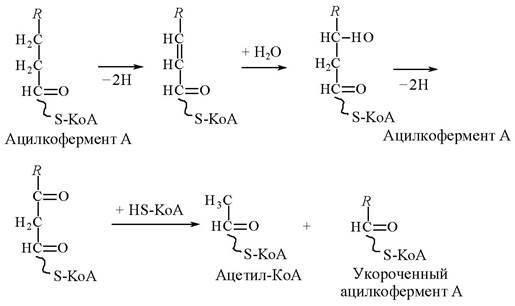
β — Окисление многократно повторяется до тех пор, пока жирная кислота полностью не превратится в ацетилкофермент А, количество молекул которого равно половине числа атомов углерода в исходной жирной кислоте. Как уже отмечалось, каждое отщепление одной молекулы ацетил — КоА сопровождается синтезом пяти молекул АТФ в ходе тканевого дыхания.
Вторым этапом окисления жирных кислот является цикл трикарбоновых кислот (ЦТК), или цикл Кребса, в котором происходит дальнейшее окисление остатка уксусной кислоты из аце- тилкофермента А до углекислого газа и воды. При окислении одной молекулы ацетил — КоА в цикле Кребса выделяется 12 молекул АТФ.
В целом окисление жирных кислот до СО2 и Н2O дает большое количество энергии. Например, в случае окисления пальмитиновой кислоты (С15Н31СООН) цикл β -окисления протекает семь раз, в результате чего образуются 35 молекул АТФ и 8 молекул ацетилкофермента А. При дальнейшем окислении 8 молекул ацетил — КоА в цикле Кребса синтезируется еще 96 молекул АТФ. Вычтя из полученной суммы молекул АТФ одну молекулу, энергия которой была затрачена на активацию жирной кислоты, получаем окончательный результат: при окислении молекулы пальмитиновой кислоты образуется 130 молекул АТФ.
Окисление жира протекает в митохондриях при обязательном использовании молекулярного кислорода, что существенно ограничивает скорость этого процесса. Поэтому за счет окисления жиров можно обеспечить энергией только работу средней мощности, но зато очень продолжительную, так как запасы жира в организме весьма значительны.
При избыточном образовании ацетилкофермента А в печени вместо цикла Кребса происходит реакция конденсации двух молекул ацетил — КоА. В результате конденсации остатки уксусной кислоты, соединяясь попарно, превращаются в кетоновые тела, а кофермент А выделяется в свободном виде:
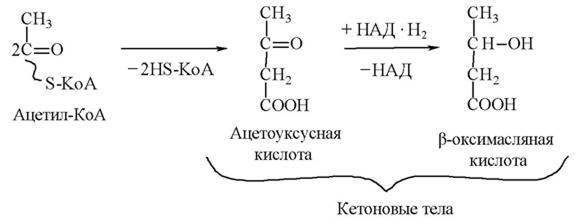
С током крови кетоновые тела поступают во все ткани.
Однако большая часть кетоновых тел извлекается из крови органами, имеющими высокие энергозатраты: миокардом, скелетными мышцами, почками.
В этих органах с участием их собственного кофермента А кетоновые тела вновь переходят в ацетилкофермент А:
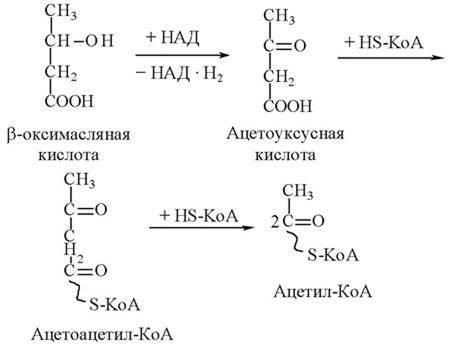
Далее ацетилкофермент А окисляется в цикле Кребса и обеспечивает эти органы энергией, необходимой для их функционирования. Особенно велика роль кетоновых тел в обеспечении организма энергией при продолжительных физических нагрузках.
При накоплении кетоновых тел в крови возможно образование ацетона:
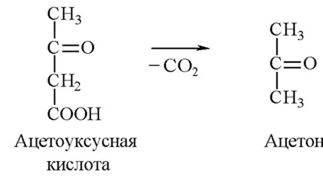
Это вещество в качестве источника энергии в организме не используется и выделяется легкими и почками.
ПредыдущаяСледующая