Синапс
Содержание:
Механизм синаптической передачи
4 этап
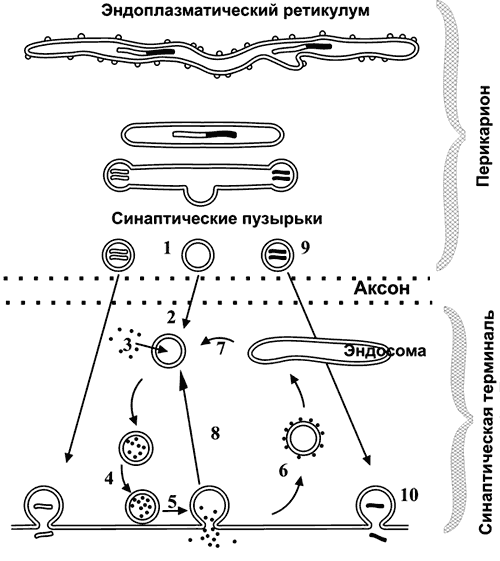
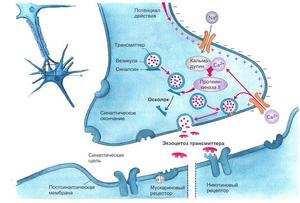
Ионы Ca вызывают образование специального белкового комплекса, который включает в себя везикулу и структуры, расположенные непосредственно около пресинаптической мембраны.
Они связаны между собой так называемыми белками экзоцитоза.
Часть белков расположена на везикулах (синапсин, синаптотагмин, синаптобревин), а часть — на пресинаптической мембране (синтаксин, синапсо-ассоциированный белок). Данный комплекс получил название секретосома.
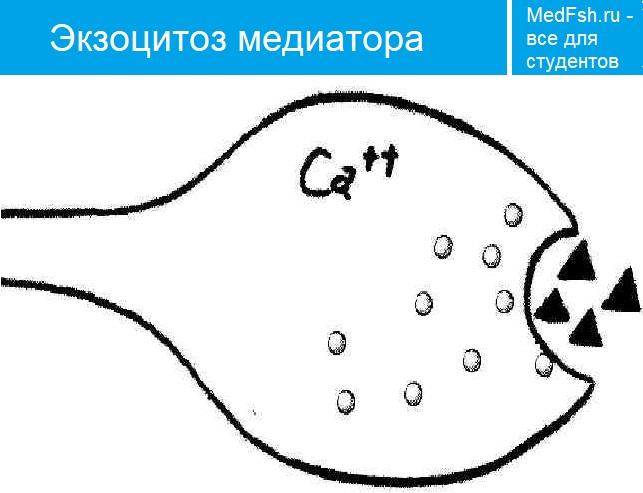
Излитию содержимого пузырька в щель способствует белок синаптопорин, формирующий канал, по которому идет выброс медиатора.
Квант медиатора — количество молекул, содержащихся в одной везикуле.
На 1 ПД выбрасывается 100 квантов АХ.
10 этап
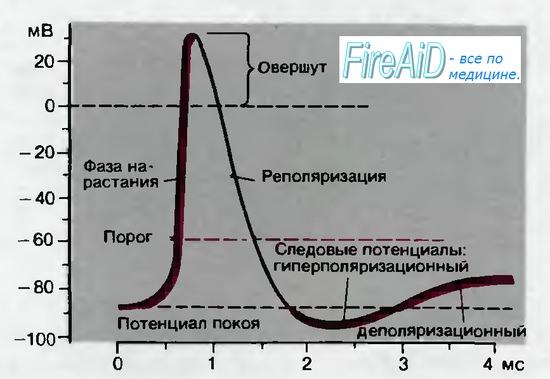
На постсинаптической мембране возникает потенциал концевой пластинки (ПКП). Он является аналогом локального ответа (ЛО).
Потенциал действия на постсинаптической мембране не возникает! Он формируется на соседней мембране мышечного волокна.
Судьба медиатора:
- связывание с рецептором,
- разрушение ферментов (ацетилхолинэстеразой),
- обратное поглощение в пресинаптическую мембрану,
- вымывание из щели и фагоцитоз.
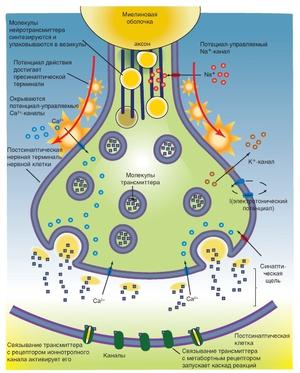
События в синапсе:
- ПД приходит к терминали аксона;
- Он деполяризует пресинаптическую мембрану;
- Ca2+ входит в терминаль, что приводит к выделению АХ;
- В синаптическую щель выделяется медиатор АХ;
- Он диффундирует в щель и связывается с рецепторами постсинаптической мембраны;
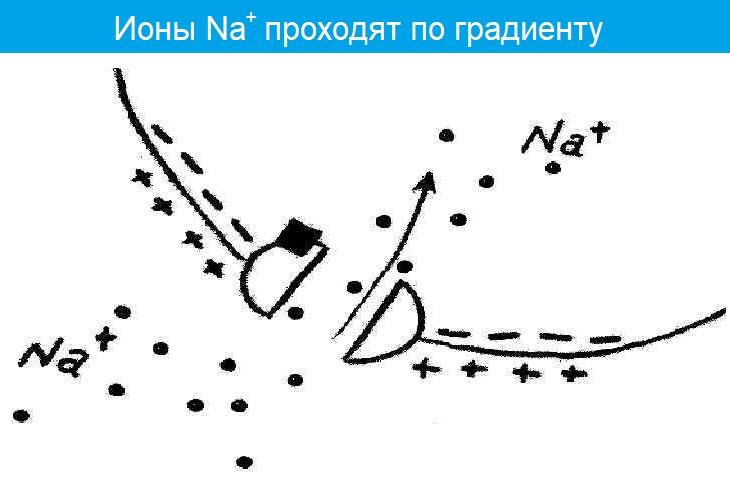
- Меняется проницаемость постсинаптической мембраны для ионов Na+;
- Ионы Na+ проникают в постсинаптическую мембрану и уменьшают ее заряд — возникает потенциал концевой пластинки (ПКП).
На самой постсинаптической мембране ПД возникнуть не может, так как здесь отсутствуют потенциалзависимые каналы, они являются хемозависимыми!
- ПКП суммируются и достигают КУД на соседнем участке мышечного волокна, что приводит к возникновению ПД и его распространению по мышечному волокну (около 5 м/с).
Достигнув пороговой величины, то есть КУД, ПКП возбуждает соседнюю (внесинаптическую) мембрану мышечного волокна за счет местных круговых токов.

Умственная работоспособность детей
 Работоспособность человека напрямую определяет его возраст, когда все значения увеличиваются одновременно с развитием и физическим ростом детей.
Работоспособность человека напрямую определяет его возраст, когда все значения увеличиваются одновременно с развитием и физическим ростом детей.
Точность и скорость умственных действий с возрастом осуществляется неравномерно в зависимости от прочих факторов, фиксирующих развитие и физический рост организма. Учащимся любого возраста, у которых присутствуют отклонения здоровья, характерна работоспособность низкого значения относительно окружающих крепких детей.
У здоровых первоклашек со сниженной готовностью организма к постоянному процессу обучения по некоторым показателям способность к действию является низкой, что усложняет борьбу с возникающими проблемами в процессе учёбы.
Скорость наступления ослабленности обуславливается исходным состоянием детской системы чувствительного нервного генеза, рабочим темпом и объёмом нагрузки. При этом дети склонны к переутомлению во время продолжительной неподвижности и когда выполняемые действия ребёнку неинтересны. После перерыва работоспособность становится прежней или становится выше прежней, причём лучше отдых делать не пассивным, но активным, переключившись на отличное от этого занятие.
Изменение работоспособности у детей
 Первая часть учебного процесса у обычных детей начальных классов сопровождается отличной работоспособностью, но к окончанию 3 урока у них отмечается снижение концентрации внимания:
Первая часть учебного процесса у обычных детей начальных классов сопровождается отличной работоспособностью, но к окончанию 3 урока у них отмечается снижение концентрации внимания:
- Они глядят в окно.
- Невнимательно слушают слова учителя.
- Изменяют положения своего тела.
- Начинают разговаривать.
- Встают со своего места.
Специфически велики значения работоспособности у старшеклассников, обучающихся во 2 смену
Особенно важно обратить внимание на то, что достаточно коротко время для подготовки к занятиям до времени начала учебного действия в классе и не гарантирует полноценного избавления от пагубных изменений в центральной нервной системе. Умственная активность быстро истощается в первые часы уроков, что явно отмечается в отрицательном поведении
Посему качественные сдвиги работоспособности наблюдаются у учеников младшего блока на уроках с 1 — 3, а блоках среднего-старшего звена на 4 — 5 занятии. В свою очередь, 6 урок проходит в условиях особенно сниженной способности к действию. При этом продолжительность занятия у 2 — 11 классников – 45 минут, что ослабляет состояние детей. Поэтому рекомендуется периодически сменять вид работы, а в середине урока провести активную паузу.
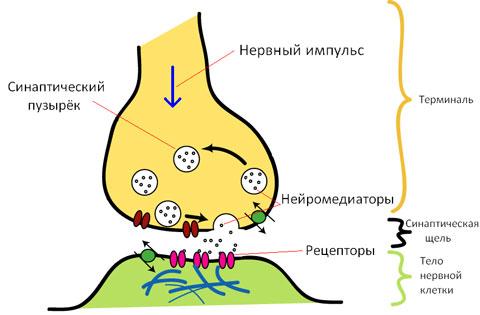
Инактивирование медиатора
Инактивирование (полная потеря активности) медиатора необходимо для реполяризации постсинаптической мембраны и восстановления исходного уровня мембранного потенциала.
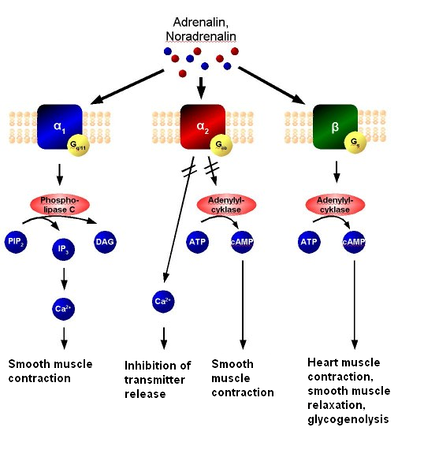
Наиболее важным путем инактивирования является ферментативный ГИДршиз медиатора с помощью ингибиторов. Для ацетилхолина ингибитором является холинэстераза. Для норадреналина и адреналина — моноаминоксидаза (МАО) и др.
В результате инактивирования медиатора возбуждение прекращается и возникает снова только при поступлении следующей порции. Возбуждение постсинаптической мембраны носит, таким образом, импульсный, дискретный, т.е. квантовый характер.
Другой путь удаления медиатора из синаптической щели — «обратный захват», обратный (ретроградный) аксонный транспорт, особенно выраженный для катехоламинов.
Классификации синапсов
Основные элементы электрического синапса (эфапса): а — коннексон в закрытом состоянии; b — коннексон в открытом состоянии; с — коннексон, встроенный в мембрану; d — мономер коннексина, е — плазматическая мембрана; f — межклеточное пространство; g — промежуток в 2-4 нанометра в электрическом синапсе; h — гидрофильный канал коннексона
По механизму передачи нервного импульса
- химический — это место близкого прилегания двух нервных клеток, для передачи нервного импульса через которое клетка-источник выпускает в межклеточное пространство особое вещество, нейромедиатор, присутствие которого в синаптической щели возбуждает или затормаживает клетку-приёмник.
- электрический (эфапс) — место более близкого прилегания пары клеток, где их мембраны соединяются с помощью особых белковых образований — коннексонов (каждый коннексон состоит из шести белковых субъединиц). Расстояние между мембранами клетки в электрическом синапсе — 3,5 нм (обычное межклеточное — 20 нм). Так как сопротивление внеклеточной жидкости мало (в данном случае), импульсы через синапс проходят не задерживаясь. Электрические синапсы обычно бывают возбуждающими.
- смешанные синапсы — пресинаптический потенциал действия создает ток, который деполяризует постсинаптическую мембрану типичного химического синапса, где пре- и постсинаптические мембраны не плотно прилегают друг к другу. Таким образом, в этих синапсах химическая передача служит необходимым усиливающим механизмом.
Наиболее распространены химические синапсы. Для нервной системы млекопитающих электрические синапсы менее характерны, чем химические.
По местоположению и принадлежности структурам
- периферические
- нервно-мышечные
- нейросекреторные (аксо-вазальные)
- рецепторно-нейрональные
- центральные
-
аксо-дендритические — с дендритами
аксо-шипиковые — с дендритными шипиками, выростами на дендритах;
, в том числе
- аксо-соматические — с телами нейронов;
- аксо-аксональные — между аксонами;
- дендро-дендритические — между дендритами;
-
аксо-дендритические — с дендритами
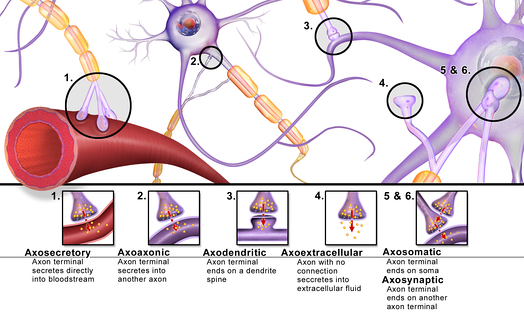
Различные варианты расположения химических синапсов
По нейромедиатору
- аминергические, содержащие биогенные амины (например, серотонин, дофамин
в том числе адренергические, содержащие адреналин или норадреналин;
);
- холинергические, содержащие ацетилхолин;
- пуринергические, содержащие пурины;
- пептидергические, содержащие пептиды.
При этом в синапсе не всегда вырабатывается только один медиатор. Обычно основной медиатор выбрасывается вместе с другим, играющим роль модулятора.
По знаку действия синапса
- возбуждающие
- тормозные.
Если первые способствуют возникновению возбуждения в постсинаптической клетке (в них в результате поступления импульса происходит деполяризация мембраны, которая может вызвать потенциал действия при определённых условиях), то вторые, напротив, прекращают или предотвращают его появление, препятствуют дальнейшему распространению импульса. Обычно тормозными являются глицинергические (медиатор — глицин) и ГАМК-ергические синапсы (медиатор — гамма-аминомасляная кислота).
Тормозные синапсы бывают двух видов:
1) синапс, в пресинаптических окончаниях которого выделяется медиатор, гиперполяризующий постсинаптическую мембрану и вызывающий возникновение тормозного постсинаптического потенциала;
2) аксо-аксональный синапс, обеспечивающий пресинаптическое торможение.
В некоторых синапсах присутствует постсинаптическое уплотнение — электронно-плотная зона, состоящая из белков. По её наличию или отсутствию выделяют синапсы асимметричные и симметричные. Известно, что все глутаматергические синапсы асимметричны, а ГАМКергические — симметричны.
В случаях, когда с постсинаптической мембраной контактирует несколько синаптических расширений, образуются множественные синапсы.
К специальным формам синапсов относятся шипиковые аппараты, в которых с синаптическим расширением контактируют короткие одиночные или множественные выпячивания постсинаптической мембраны дендрита. Шипиковые аппараты значительно увеличивают количество синаптических контактов на нейроне и, следовательно, количество перерабатываемой информации. «Не-шипиковые» синапсы называются «сидячими». Например, сидячими являются все ГАМК-ергические синапсы.
История открытия
- В 1897 году Шеррингтон сформулировал представление о синапсах.
- За исследования нервной системы, в том числе синаптической передачи, в 1906 году Нобелевскую премию получили Гольджи и Рамон-и-Кахаль.
- В 1921 австрийский учёный О. Лёви (О. Loewi) установил химическую природу передачи возбуждения через синапсы и роль в ней ацетилхолина. Получил Нобелевскую премию в 1936 г. совместно с Г. Дейлом (Н. Dale).
- В 1933 советский учёный А. В. Кибяков установил роль адреналина в синаптической передаче.
- 1970 — Б. Кац (В. Katz, Великобритания), У. фон Эйлер (U. v. Euler, Швеция) и Дж. Аксельрод (J. Axelrod, США) получили Нобелевскую премию за открытие роли норадреналина в синаптической передаче.
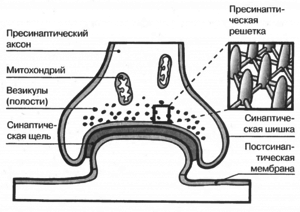
Строение синапса
Синапс состоит из трех основных элементов:
- Пресинаптической мембраны, которая покрывает расширенное нервное окончание, представляющее собой нейросекреторный аппарат. В пресинаптической части находятся пузырьки и митохондрии, обеспечивающие синтез медиатора. Медиаторы депонируются в гранулах (пузырьках).
- Постсинаптической мембраны – это утолщенная часть мембраны клетки, с которой контактирует пресинаптическая мембрана. Она имеет ионные каналы и способна к генерации потенциала действия. Кроме того, на ней расположены специальные белковые структуры — рецепторы, воспринимающие действие медиаторов.
- Синаптической щели, она представляет собой пространство между пресинаптической и постсинаптической мембранами, заполненное жидкостью, близкой по составу к плазме крови.
Вот так строение синапса выглядит на рисунке.
Синапсы электрического и химического направления
 Действие слияния плёнок-мембран принято считать электрическими синапсами. В условиях, когда необходимая синаптическая щель прерывистая с промежутками перегородок монолитного соединения. Эти перегородки формируют чередующуюся конструкцию отделений синапса, при этом отделения обособлены фрагментами приближенных мембран, промежуток между которыми в синапсах обычного склада равен 0,15 — 0,20 нм у представителей млекопитающих существ. В месте соединения плёнок-мембран присутствуют пути, с помощью которых происходит обмен частью плодов.
Действие слияния плёнок-мембран принято считать электрическими синапсами. В условиях, когда необходимая синаптическая щель прерывистая с промежутками перегородок монолитного соединения. Эти перегородки формируют чередующуюся конструкцию отделений синапса, при этом отделения обособлены фрагментами приближенных мембран, промежуток между которыми в синапсах обычного склада равен 0,15 — 0,20 нм у представителей млекопитающих существ. В месте соединения плёнок-мембран присутствуют пути, с помощью которых происходит обмен частью плодов.
Помимо отдельчатых типов синапсов существуют необходимые электрические типичные синапсы в виде единой синаптической щели, общий периметр которой простирается на 1000 мкм. Так, подобное синаптическое явление представлено в нейронах ресничного ганглия.
Электрические синапсы способны проводить качественное возбуждение в одностороннем порядке. Этот факт отмечается при фиксации электрического резерва синаптической составляющей. Например, в момент при касании афферентных канальцев синаптическая плёнка-мембрана деполяризуется, когда с касанием эфферентных частиц волокон напортив — гиперполяризуется. Считается, что синапсы действующих нейронов с общими обязанностями могут осуществлять требуемое возбуждение (между 2 пропускающими участками) в обе стороны.
Напротив, синапсы присутствующих нейронов с разным перечнем действий (моторные и сенсорные) проводят акт возбуждения односторонне. Основная работа синаптических составляющих обуславливается продуцированием безотлагательных реакций организма. Электрический синапс подлежит незначительной доли утомляемости, обладает значительным процентом устойчивости к внутренне-наружным факторам.
Химические синапсы обладают видом предсинаптического сегмента, функциональной синаптической щели с фрагментом постсинаптической составляющей. Предсинаптический фрагмент формируется увеличением размера аксона внутри собственного канальца или к его завершению. В этом фрагменте присутствуют гранулярные, а также агранулярные специальные мешочки, содержащие медиатор.
Предсинаптическое увеличение наблюдает локализацию активных митохондрий, генерирующую частицы вещества-гликогена, а также требуемую выработку медиатора и другое. В условиях частого соприкосновения с предсинаптическим полем резерв медиатора в имеющихся мешочках утрачивается.
Существует мнение, что малые гранулярные пузырьки имеют такое вещество, как норадреналин, а большие – катехоламины. Причём в агранулярных полостях (пузырьках) располагается ацетилхонин. Помимо этого, медиаторами усиленного возбуждения считаются вещества, образованные по типу вырабатываемой аспарагиновой или не менее значимой кислоты глутамина.
Действующие контакты синапса часто располагаются между:
- Дендритом и аксоном.
- Сомой и аксоном.
- Дендритами.
- Аксонами.
- Сомой клетки и дендритами.
Влияние выработанного медиатора относительно присутствующей постсинаптической плёнки-мембраны происходит из-за чрезмерного проникновения её частиц натрия. Генерация мощных изливаний частиц натрия из рабочей синаптической щели сквозь постсинаптическую плёнку-мембрану формирует её деполяризацию, образуя возбуждение постсинаптического резерва. Транзиту химического направления данных синапса свойственно синаптическое приостановление возбуждения по времени равному 0,5 мс с выработкой постсинаптического резерва, как реакция на предсинаптический поток.
Данная возможность в момент возбуждения представляется в деполяризации постсинаптической плёнки-мембраны, а в момент приостановления в её гиперполяризации. Из-за чего наблюдается приостановленный постсинаптический резерв. Как правило, во время сильного возбуждения повышается уровень проницаемости постсинаптической плёнки-мембраны.
Требуемое возбуждающее свойство фиксируется внутри нейронов, если в типичных синапсах работает норадреналин, вещество-дофамин, ацетил холин, важный серотонин, вещество Р и кислота глутамина.
Сдерживающий потенциал формируется во время влияния на синапсы из гамма-аминомаслянной кислоты и глицина.