Потенциал покоя
Содержание:
- Активные свойства мембраны
- Потенциал действия нервного волокна, нервный импульс
- История
- Общие положения
- Общие положения Править
- Распространение потенциала действия между клетками
- Рефрактерные периоды и физиология образования потенциала действия клетки
- Распространение потенциала действия Править
- Физиологические свойства возбудимых тканей
- Потенциал действия. Временной ход потенциала действия. Реполяризация.
- Уравнения Голдмана
- Транспорт веществ
- Как работают потенциалы действия?
- Потенциал действия в различных типах клеток
Активные свойства мембраны
Схема строения мембраны клетки.
Активные свойства мембраны, обеспечивающие возникновение потенциала действия, основываются главным образом на поведении потенциалзависимых натриевых (Na+-) и калиевых (K+-) каналов. Начальная фаза ПД формируется входящим натриевым током, позже открываются калиевые каналы и выходящий K+-ток возвращает потенциал мембраны к исходному уровню. Исходную концентрацию ионов затем восстанавливает натрий-калиевый насос.
По ходу ПД каналы переходят из состояния в состояние: у Na+-каналов основных состояний три — закрытое, открытое и инактивированное (в реальности дело сложнее, но этих трёх достаточно для описания), у K+-каналов два — закрытое и открытое.
Поведение каналов, участвующих в формировании ПД, описывается через проводимость и вычисляется через коэффициенты переноса (трансфера).
Коэффициенты переноса были выведены Ходжкином и Хаксли.
- Проводимость для калия GK на единицу площади [S/cm²]
| dndt=αn(1−n)−βnn{\displaystyle dn/dt=\alpha _{n}(1-n)-\beta _{n}n}, |
| где: |
| αn{\displaystyle \alpha _{n}} — коэффициент трансфера из закрытого в открытое состояние для K+-каналов [1/s]; |
| βn{\displaystyle \beta _{n}} — коэффициент трансфера из открытого в закрытое состояние для K+-каналов [1/s]; |
| n{\displaystyle n} — фракция K+-каналов в открытом состоянии; |
| (1−n){\displaystyle (1-n)} — фракция K+-каналов в закрытом состоянии |
- Проводимость для натрия GNa на единицу площади [S/cm²]
рассчитывается сложнее, поскольку, как уже было сказано, у потенциал-зависимых Na+-каналов, помимо закрытого/открытого состояний, переход между которыми описывается параметром m{\displaystyle m}, есть ещё инактивированное/не-инактивированное состояния, переход между которыми описывается через параметр h{\displaystyle h}
| dmdt=αm(1−m)−βmm{\displaystyle dm/dt=\alpha _{m}(1-m)-\beta _{m}m}, | dhdt=αh(1−h)−βhh{\displaystyle dh/dt=\alpha _{h}(1-h)-\beta _{h}h}, |
| где: | где: |
| αm{\displaystyle \alpha _{m}} — коэффициент трансфера из закрытого в открытое состояние для Na+-каналов [1/s]; | αh{\displaystyle \alpha _{h}} — коэффициент трансфера из инактивированного в не-инактивированное состояние для Na+-каналов [1/s]; |
| βm{\displaystyle \beta _{m}} — коэффициент трансфера из открытого в закрытое состояние для Na+-каналов [1/s]; | βh{\displaystyle \beta _{h}} — коэффициент трансфера из не-инактивированного в инактивированное состояние для Na+-каналов [1/s]; |
| m{\displaystyle m} — фракция Na+-каналов в открытом состоянии; | h{\displaystyle h} — фракция Na+-каналов в не-инактивированном состоянии; |
| (1−m){\displaystyle (1-m)} — фракция Na+-каналов в закрытом состоянии | (1−h){\displaystyle (1-h)} — фракция Na+-каналов в инактивированном состоянии. |
Потенциал действия нервного волокна, нервный импульс
Потенциал действия является основным специфическим признаком возбуждения. Его регистрация свидетельствует о том, что клетка или ее структуры ответили на воздействие возбуждением. Однако, как уже отмечалось, ПД в некоторых клетках может возникать спонтанно (самопроизвольно). Такие клетки содержатся в водителях ритма сердца, стенках сосудов, нервной системе. ПД используется как носитель информации, передающий ее в виде электрических сигналов (электрическая сигнализации) по афферентным и эфферентным нервным волокнам, проводящей системе сердца, а также для инициирования сокращения мышечных клеток.
Рассмотрим причины и механизм генерации ПД в афферентных нервных волокнах, образующих первично воспринимающие сенсорные рецепторы. Непосредственной причиной возникновения (генерации) ПД в них является рецепторный потенциал.
Если измерять разность потенциалов на мембране ближайшего к нервному окончанию перехвата Ранвье, то в промежутках между воздействиями на капсулу тельца Пачини она остается неизменной (70 мВ), а во время воздействия деполяризуется почти одновременно с деполяризацией рецепторной мембраны нервного окончания.
При увеличении силы давления на тельце Пачини, вызывающей возрастание рецепторного потенциала до 10 мВ, в ближайшем перехвате Ранвье обычно регистрируется быстрое колебание мембранного потенциала, сопровождающееся перезарядкой мембраны — потенциал действия (ПД), или нервный импульс (рис. 2). Если сила давления на тельце возрастет еще больше, амплитуда рецепторного потенциала увеличивается и в нервном окончании генерируется уже ряд потенциалов действия с определенной частотой.
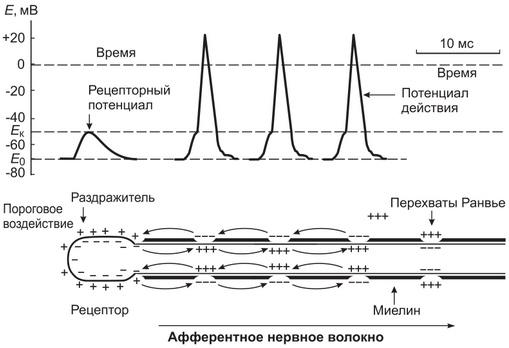
Рис. 2. Схематическое представление механизма преобразования рецепторного потенциала в потенциал действия (нервный импульс) и распространения импульса по нервному волокну
Суть механизма генерации ПД состоит в том, что рецепторный потенциал вызывает возникновение локальных круговых токов между деполяризованной рецепторной мембраной немиелинизированной части нервного окончания и мембраной первого перехвата Ранвье. Эти токи, носителями которых являются ионы Na+, К+, СI- и другие минеральные ионы, «протекают» не только вдоль, но и поперек мембраны нервного волокна в области перехвата Ранвье. В мембране перехватов Ранвье в отличие от рецепторной мембраны самого нервного окончания имеется большая плотность ионных потенциалзависимых натриевых и калиевых каналов.
При достижении на мембране перехвата Ранвье величины деполяризации около 10 мВ происходит открытие быстрых потенциалзависимых натриевых каналов и через них в аксоплазму по электрохимическому градиенту устремляется поток ионов Na+. Он обусловливает быструю деполяризацию и перезарядку мембраны перехвата Ранвье. Однако одновременно с открытием быстрых потенциалзависимых натриевых каналов в мембране перехвата Ранвье открываются медленные потенциалзависимые калиевые каналы и из аксоилазмы начинают выходить ионы К+ Их выход запаздывает по отношению ко входу ионов Na+. Таким образом, входящие с большой скоростью в аксоплазму ионы Na+ быстро деполяризуют и перезаряжают на короткое время (0,3-0,5 мс) мембрану, а выходящие ионы К+ восстанавливают исходное распределение зарядов на мембране (реполяризуют мембрану). В результате во время механического воздействия на тельце Пачини силой, равной или превышающей пороговую, на мембране ближайшего перехвата Ранвье наблюдается кратковременное колебание потенциала в виде быстрой деполяризации и реполяризации мембраны, т.е. генерируется ПД (нервный импульс).
Поскольку непосредственной причиной генерации ПД является рецепторный потенциал, то его в этом случае еще называют генераторным потенциалом. Число генерируемых в единицу времени одинаковых по амплитуде и длительности нервных импульсов пропорционально амплитуде рецепторного потенциала, а следовательно, силе давления на рецептор. Процесс преобразования информации о силе воздействия, заложенной в амплитуде рецепторного потенциала, в число дискретных нервных импульсов получил название дискретного кодирования информации.
Более подробно ионные механизмы и временная динамика процессов генерации ПД изучены в экспериментальных условиях при искусственном воздействии на нервное волокно электрическим током различной силы и длительности.
История
Основные положения мембранной теории возбуждения сформулированы немецким нейрофизиологом Ю.Бернштейном
В 1902 году Юлиус Бернштейн выдвинул гипотезу, согласно которой клеточная мембрана пропускает внутрь клетки ионы К +, и они накапливаются в цитоплазме. Расчет величины потенциала покоя по уравнению Нернста для калиевого электрода удовлетворительно совпал с измеренным потенциалом между саркоплазме мышцы и окружающей средой, который составил около — 70 мВ. Согласно теории Ю.Бернштейна, при возбуждении клетки ее мембрана повреждается, и ионы К + выходят из клетки по концентрационном градиента до тех пор, пока потенциал мембраны не становится равным нулю. Затем мембрана восстанавливает свою целостность, и потенциал возвращается к уровню потенциала покоя.
Эту модель развили в своей работе 1952 Алан Ллойд Ходжкин и Эндрю Хаксли в которой описали электрические механизмы, обусловливающие генерацию и передачу нервного сигнала в гигантском аксоне кальмара. За это авторы модели получили Нобелевскую премию в области физиологии и медицины за 1963 год. Модель получила название модель Ходжкина-Хаксли
В 2005 году Томасом Геймбургом и Анрю Д. Джексоном предложена солитонном модель, основанная на предположении, что сигнал по нейронам распространяется в виде солитонов — устойчивых волн, распространяющихся по клеточной мембране.
Общие положения
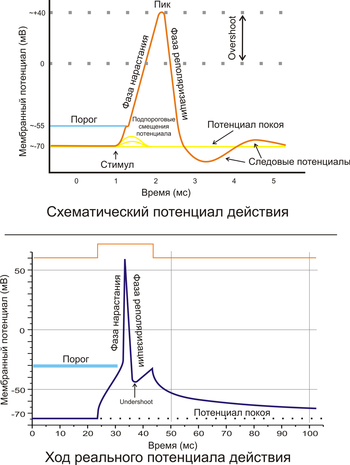 Рис. 2. A. Схематичное изображение идеализированного потенциала действия. B. Реальный потенциал действия пирамидного нейрона гиппокампа крысы. Форма реального потенциала действия обычно отличается от идеализированной.
Рис. 2. A. Схематичное изображение идеализированного потенциала действия. B. Реальный потенциал действия пирамидного нейрона гиппокампа крысы. Форма реального потенциала действия обычно отличается от идеализированной.
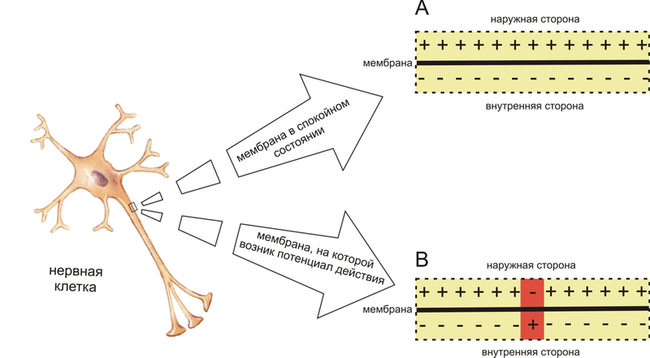
Поляризация мембраны живой клетки обусловлена отличием ионного состава с её внутренней и наружной стороны. Когда клетка находится в спокойном (невозбуждённом) состоянии, ионы по разные стороны мембраны создают относительно стабильную разность потенциалов, называемую потенциалом покоя. Если ввести внутрь живой клетки электрод и измерить мембранный потенциал покоя, он будет иметь отрицательное значение (около −70 — −90 мВ). Это объясняется тем, что суммарный заряд на внутренней стороне мембраны существенно меньше, чем на внешней, хотя с обеих сторон содержатся и катионы, и анионы. Снаружи — на порядок больше ионов натрия, кальция и хлора, внутри — ионов калия и отрицательно заряженных белковых молекул, аминокислот, органических кислот, фосфатов, сульфатов. Надо понимать, что речь идёт именно о заряде поверхности мембраны — в целом среда и внутри, и снаружи клетки заряжена нейтрально.
Потенциал мембраны может изменяться под действием различных стимулов. Искусственным стимулом может служить электрический ток, подаваемый на внешнюю или внутреннюю сторону мембраны через электрод. В естественных условиях стимулом часто служит химический сигнал от соседних клеток, поступающий через синапс или путём диффузной передачи через межклеточную среду. Смещение мембранного потенциала может происходить в отрицательную (гиперполяризация) или положительную (деполяризация) сторону.
В нервной ткани потенциал действия, как правило, возникает при деполяризации — если деполяризация мембраны нейрона достигает некоторого порогового уровня или превышает его, клетка возбуждается, и от её тела к аксонам и дендритам распространяется волна электрического сигнала. (В реальных условиях на теле нейрона обычно возникают постсинаптические потенциалы, которые сильно отличаются от потенциала действия по своей природе — например, они не подчиняются принципу «всё или ничего». Эти потенциалы преобразуются в потенциал действия на особом участке мембраны — аксонном холмике, так что потенциал действия не распространяется на дендриты).
Рис. 3. Простейшая схема, демонстрирующая мембрану с двумя натриевыми каналами в открытом и закрытом состоянии
Это обусловлено тем, что на мембране клетки находятся ионные каналы — белковые молекулы, образующие в мембране поры, через которые ионы могут проходить с внутренней стороны мембраны на наружную и наоборот. Большинство каналов ионо-специфичны — натриевый канал пропускает практически только ионы натрия и не пропускает другие (это явление называют селективностью). Мембрана клеток возбудимых тканей (нервной и мышечной) содержит большое количество потенциал-зависимых ионных каналов, способных быстро реагировать на смещение мембранного потенциала. Деполяризация мембраны в первую очередь вызывает открытие потенциал-зависимых натриевых каналов. Когда одновременно открывается достаточно много натриевых каналов, положительно заряженные ионы натрия устремляются через них на внутреннюю сторону мембраны. Движущая сила в данном случае обеспечивается градиентом концентрации (с внешней стороны мембраны находится намного больше положительно заряженных ионов натрия, чем внутри клетки) и отрицательным зарядом внутренней стороны мембраны (см. Рис. 2).
Поток ионов натрия вызывает ещё бо́льшее и очень быстрое изменение мембранного потенциала, которое и называют потенциалом действия (в специальной литературе обозначается ПД).
Согласно закону «всё-или-ничего» мембрана клетки возбудимой ткани либо не отвечает на стимул совсем, либо отвечает с максимально возможной для неё на данный момент силой. То есть, если стимул слишком слаб и порог не достигнут, потенциал действия не возникает совсем; в то же время, пороговый стимул вызовет потенциал действия такой же амплитуды, как и стимул, превышающий пороговый. Это отнюдь не означает, что амплитуда потенциала действия всегда одинакова — один и тот же участок мембраны, находясь в разных состояниях, может генерировать потенциалы действия разной амплитуды.
После возбуждения нейрон на некоторое время оказывается в состоянии абсолютной рефрактерности, когда никакие сигналы не могут его возбудить снова, затем входит в фазу относительной рефрактерности, когда его могут возбудить исключительно сильные сигналы (при этом амплитуда ПД будет ниже, чем обычно). Рефрактерный период возникает из-за инактивации быстрого натриевого тока, то есть инактивации натриевых каналов (см. ниже).
Общие положения Править
Для того, чтобы на мембране поддерживалась разность потенциалов, необходимо, чтобы была определенная разность концентрации различных ионов внутри и снаружи клетки.
Концентрации ионов в клетке скелетной мышцы и во внеклеточной среде
| Ионы | Концентрация в саркоплазме (ммоль) | Концентрация вне клетки (ммоль) |
|---|---|---|
| K+ | 140 | 2,5 |
| Na+ | 10 | 120 |
| Cl- | 3-4 | 120 |
| Ca2+ | <0,001 | 2 |
| A- (полипептиды) | 140 |
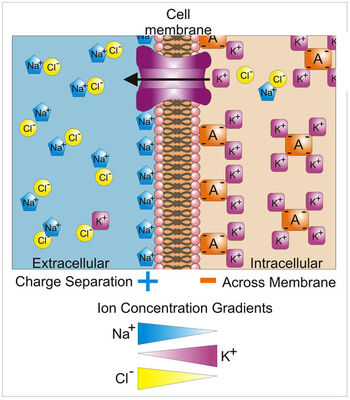
Различия в концентрации ионов на противоположных сторонах клеточной мембраны приводят к напряжению, названному мембранным потенциалом. Много ионов имеют градиент концентрации поперек мембраны, включая калий (K+), который является в высокой внутренней части и низкой концентрации вне мембраны. Натрий (Na+) и хлорид (Замкнутый–) ионы — при высоких концентрациях во внеклеточной области, и низких концентрациях во внутриклеточных областях. Эти градиенты концентрации обеспечивают потенциальную энергию вести формирование мембранного потенциала. Это напряжение установлено, когда мембрана имеет проходимость к одному или более ионам. В самом простом случае, иллюстрированном здесь, если мембрана выборочно водопроницаема к калию, они положительно обвиняли, что ионы могут распространить вниз градиент концентрации к внешней стороне ячейки, оставляя позади неданные компенсацию отрицательные обвинения. Это разделение обвинений-, каков вызывает мембранный потенциал. Отметьте, что оптовые решения любой стороны мембраны electo-нейтральны. Аналогично, система в целом нейтральна гальваностереотипом. «Неданные компенсацию» положительные обвинения вне ячейки, и неданных компенсацию отрицательных обвинений в ячейке, физически выстраиваются в линию на мембранной поверхности и привлекают друг друга поперек мембраны. Таким образом, мембранный потенциал физически расположен только в непосредственной близости мембраны. Это — разделение этих обвинений поперек них мембрана, которая является основанием мембранного напряжения. Отметьте также, что эта диаграмма — только приближение ионных вкладов в мембранный потенциал. Другие ионы, включая натрий, хлорид, кальций и другие играют более незначительную роль, даже при том, что они имеют сильные градиенты концентрации, потому что они более ограничили проходимость чем калий. Ключ: Синие пятиугольники — ионы натрия; Фиолетовые площади — ионы калия; Желтые круги — ионы Choloride; Оранжевые прямоугольники — Анионы (они являются результатом разнообразия источников, включая белки). Большая фиолетовая структура со стрелкой представляет трансмембранный канал калия и руководство чистого движения калия.
Потенциал покоя для большинства нейронов составляет величину порядка −60 мВ — −70 мВ. У клеток невозбудимых тканей на мембране также имеется разность потенциалов, разная для клеток разных тканей и организмов.
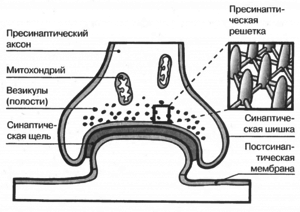
Распространение потенциала действия между клетками
В химическом синапсе после того, как волна потенциала действия доходит нервного окончания, она вызывает высвобождение нейротрансмиттеров из пресинаптических пузырьков в синаптическую щель. Молекулы медиатора, высвобождаемых с пресинапса, связываются с рецепторами на постсинаптической мембране, в результате чего в рецепторных макромолекулах открываются ионные каналы. Ионы, начинают поступать внутрь постсинаптической клетки через открытые каналы, изменяют заряд ее мембраны, что приводит к частичной деполяризации мембраны и, как следствие, провоцирование генерации постсинаптической клетки потенциала действия.
В электрическом синапсе отсутствует «посредник» передачи в виде нейромедиатора. Зато клетки соединены между собой с помощью специфических протеиновых тоннелей — конексонив, поэтому ионные токи, с пресинаптической клетки могут стимулировать постсинаптическую клетку, вызывая зарождения в ней потенциала действия. Благодаря такому строению, потенциал действия может распространяться в обе стороны и значительно быстрее, чем через химический синапс.
-
Схема процесса передачи нервного сигнала в химическом синапсе
-
Схема строения электрического синапса
Рефрактерные периоды и физиология образования потенциала действия клетки
Потенциалы действия работают на основе «все или ничего». Это означает, что потенциал действия либо срабатывает, либо не похож на щелчок переключателя. Нейрон всегда посылает потенциал действия одинакового размера. Итак, как мы можем показать, что некоторая информация более важна или требует нашего внимания прямо сейчас? Ответ заключается в том, как часто посылаются потенциалы действия — частота потенциала действия.
Когда мозг действительно возбуждается, он запускает много сигналов. Как быстро эти сигналы запускают огонь, говорит нам, насколько сильный исходный стимул — чем сильнее сигнал, тем выше частота потенциалов действия. Существует максимальная частота, с которой один нейрон может посылать потенциалы действия, и это определяется его рефрактерными периодами.
Абсолютный рефрактерный период: за это время абсолютно невозможно отправить другой потенциал действия. Ворота инактивации (h) натриевых каналов на какое-то время закрываются и делают так, что натрий не пройдет. Отсутствие натрия означает отсутствие деполяризации, что означает отсутствие потенциала действия. Абсолютные рефрактерные периоды помогают направить потенциал действия вниз по аксону, потому что только каналы дальше по течению могут открывать и впускать деполяризующие ионы.
Относительный рефрактерный период: в это время действительно сложно направить потенциал действия. Это период после абсолютного рефрактерного периода, когда ворота снова открыты. Тем не менее, ячейка по-прежнему гиперполяризована после отправки потенциала действия. Для достижения соответствующего деполяризационного потенциала потребовалось бы даже больше положительных ионов, чем обычно, чем обычно. Это означает, что исходное инициирующее событие должно быть больше нормального, чтобы передавать больше потенциалов действия. Относительные рефрактерные периоды могут помочь нам понять, насколько интенсивным является стимул — клетки в вашей сетчатке будут посылать сигналы быстрее при ярком свете, чем при тусклом свете, потому что триггер сильнее.
Периоды рефрактерности в физиологии формирования потенциалов действия также дают нейрону некоторое время для пополнения пакетов нейротрансмиттера, обнаруженного на терминале аксона, чтобы он мог продолжать передавать сообщение. Несмотря на то, что все еще возможно полностью исчерпать запас нейронов, передаваемых нейронами, путем непрерывного запуска, рефрактерные периоды помогают клетке продержаться немного дольше.
Распространение потенциала действия Править
Распространение потенциала действия по немиелинизированным волокнам Править
Если внутриклеточным электродом раздражать аксон посередине, то ПД будет распространяться в обоих направлениях. Обычно же ПД распространяется по аксону в одном направлении (от тела нейрона к нервным окончаниям), хотя деполяризация мембраны происходит по обе стороны от участка, где в данный момент возник ПД. Одностороннее проведение ПД обеспечивается свойствами натриевых каналов — после открывания они на некоторое время инактивируются и не могут открыться ни при каких значениях мембранного потенциала (свойство рефрактерности). Поэтому на ближнем к телу клетки участке, где до этого уже «прошел» ПД, он не возникает.
Распространение потенциала действия по миелинизированным волокнам Править
Распространение мембранного потенциала
сальтаторное проведение++
Чтобы представить, насколько эффективно может быть увеличена скорость проведения за счёт миелиновой оболочки, достаточно сравнить скорость распространения импульса по немиелинизированным и миелинизированным участкам нервной системы человека. При диаметре волокна около 2 µм и отсутствии миелиновой оболочки скорость проведения будет составлять ~1 м/с, а при наличии даже слабой миелинизации при том же диаметре волокна — 15-20 м/с. В волокнах большего диаметра, обладающих толстой миелинововой оболочкой, скорость проведения может достигать 120 м/с.
Следует однако понимать, что скорость распространения потенциала действия по мембране отдельно взятого нервного волокна отнюдь не является постоянной величиной — в зависимости от различных условий, эта скорость может очень значительно уменьшаться и, соответственно, увеличиваться, возвращаясь к некоему исходному уровню.
Физиологические свойства возбудимых тканей
Основными свойствами всех возбудимых тканей являются:
- возбудимость,
- проводимость,
- лабильность.
Лабильность (функциональная подвижность) — это способность ткани реагировать на различные воздействия с определенной скоростью, то есть способность воспроизводить определенную частоту раздражения.
Мерой лабильности является наибольшее число ответных реакций, которыми возбудимая ткань способна реагировать за единицу времени в соответствии с частотой приложенных к ней раздражений.
Проводимость — это способность ткани и клетки передавать возбуждение.
Раздражение — это процесс воздействия на живую ткань агентов внешней по отношению к этой ткани среды.
Раздражитель — это причина, способная вызвать ответную реакцию.
Потенциал действия. Временной ход потенциала действия. Реполяризация.
Функция нервных клеток в организме заключается в получении информации, передаче ее в другие отделы нервной системы, сопоставление информации от разных источников и, наконец, регуляции деятельности других клеток. Сигналы, поступающие от нервов, вызывают сокращение мышечных клеток. Когда эти два типа клеток «активны» (каждая по-своему), возникает быстрый сдвиг мембранного потенциала в положительном направлении — потенциал действия.
Временной ход потенциала действия
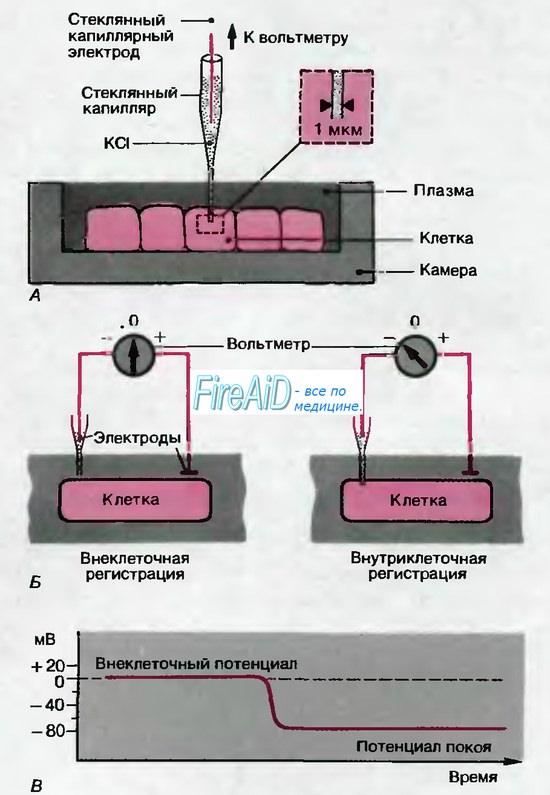
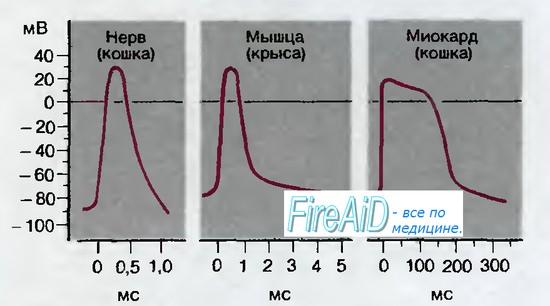
Потенциалы действия можно зарегистрировать в нервных и мышечных клетках с помощью внутриклеточных электродов (рис. 2.1). Типичные примеры потенциалов действия в различных тканях млекопитающих представлены на рис. 2.4. Во всех этих случаях потенциал резко нарастает от отрицательных значений потенциала покоя до положительного пика, составляющего примерно +30 мВ. Затем потенциал с различной скоростью возвращается к уровню покоя; длительность потенциала действия составляет около 1 мс в нервах, 10 мс в скелетной мышце и более 200 мс в миокарде.
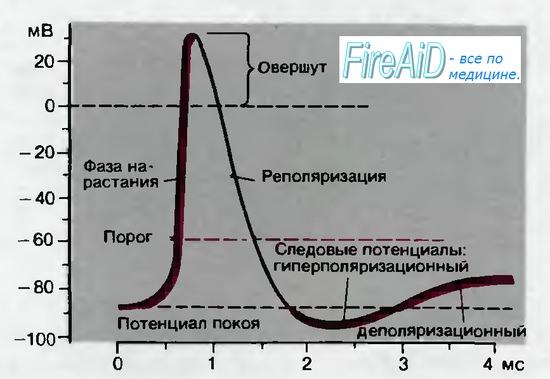
Как показывает рис. 2.5, для потенциала действия характерны несколько фаз. Он начинается очень быстрым сдвигом потенциала в положительном направлении — фазой нарастания, которая продолжается всего лишь 0,2-0,5 мс. Во время фазы нарастания клеточная мембрана теряет свой нормальный заряд («поляризацию»); поэтому фазу нарастания называют также фазой деполяризации. Обычно кривая деполяризации переходит за нулевую линию и мембранный потенциал становится положительным. Эта положительная фаза потенциала действия называется овершутом («перелетом»). Следующая за овершутом фаза, в течение которой восстанавливается исходный потенциал покоя мембраны, называется реполяризацией.
Уравнения Голдмана
Хотя равновесный потенциал для ионов калия составляет -90 мВ, МПС нейрона несколько менее отрицательный. Эта разница отражает незначительное но постоянное следование ионов Na + через мембрану в состоянии покоя. Поскольку концентрационный градиент для натрия противоположный такого для калия, Na + движется внутрь клетки и сдвигает суммарный заряд на внутренней стороне мембраны в положительную сторону. На самом деле МПС нейрона составляет от -60 до -80 мВ. Это значение значительно ближе к Е K чем до Е Na, потому что в состоянии покоя в нейроне открыто много калиевых каналов и очень мало натриевых. Также на встанвлення МПС влияет движение ионов хлора. В 1943 году Дэвид Голдаман предложил усовершенствовать уравнение Нернста так, чтобы оно отражало влияние различных ионов на мембарнний потенциал, в этом уравнении учитывается относительная проницаемость мембраны для каждого типа ионов:
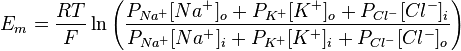
где R — универсальная газовая постоянная, Т — абслоютна температура (по шкале Кельвина), z — заряд иона, F — число Фарадея, o,i — концентрации ионов внутри и внутри клеток, Р — относительная проницаемость мембраны для соответствующего иона. Значение заряда в данном уравнении не сохраняется, но оно учтено — для хлора внешняя и внутренняя концентрация поменяны местами, так как его заряд 1.
Транспорт веществ
Транспорт частиц через каналы является жизненно важным для клеток процессом.
Обычно транспорт веществ делят на пассивный (без затраты энергии), т.е. транспорт веществ по концентрационному, осмотическому и электрохимическому градиентам и активный (с затратой энергии).
Различают первично- и вторично-активный транспорт
Первично-активный транспорт ионов обеспечивается специальными ионными насосами, осуществляется с затратой энергии АТФ, против концентрационного градиента, т.е. перенос веществ происходит из меньшей концентрации через мембрану в большую концентрацию.
Вторично-активный транспорт:
- Это вид транспорта для переноса веществ (глюкозы, аминокислот и др.) через мембрану также против градиента, но без затраты энергии.
- Эти вещества проходят через мембрану с помощью специальных переносчиков (напр. ионов Na), на транспорт которых и затрачивается энергия, а эти вещества движутся как бы попутно.
Как работают потенциалы действия?
Потенциалы действия (те электрические импульсы, которые посылают сигналы вокруг вашего тела) являются не чем иным, как временным сдвигом (с отрицательного на положительный) мембранного потенциала нейрона, вызванного ионами, внезапно входящими и выходящими из нейрона. В состоянии покоя (до возникновения потенциала действия) все закрытые натриевые и калиевые каналы закрыты. Эти закрытые каналы отличаются от каналов утечки и открываются только после срабатывания потенциала действия. Мы говорим, что эти каналы являются «управляемыми напряжением», потому что они открыты и закрыты в зависимости от разности напряжений на мембране ячейки. Натриевые каналы, управляемые напряжением, имеют два вентиля (калитка м и калитка h), а калиевый канал.
Есть три основных события, которые происходят во время действия потенциала:
Инициирующее событие, которое деполяризует тело клетки. Этот сигнал исходит от других клеток, соединяющихся с нейроном, и вызывает попадание положительно заряженных ионов в тело клетки. Положительные ионы все еще поступают в клетку, чтобы деполяризовать ее, но эти ионы проходят через каналы, которые открываются, когда конкретное химическое вещество, известное как нейротрансмиттер, связывается с каналом и приказывает ему открыться.
Нейротрансмиттеры высвобождаются клетками вблизи дендритов, часто как конечный результат их собственного потенциала действия! Эти поступающие ионы приближают потенциал мембраны к 0, что называется деполяризацией. Объект является полярным, если есть разница между более отрицательными и более положительными областями. По мере того как положительные ионы попадают в отрицательную ячейку, эта разница и, следовательно, полярность ячейки уменьшаются. Если тело ячейки становится достаточно положительным, чтобы оно могло запустить натриевые каналы с управляемым напряжением, найденные в аксоне, то потенциал действия будет послан.
Деполяризация — делает клетку менее полярной (мембранный потенциал уменьшается, когда ионы быстро начинают выравнивать градиенты концентрации). Управляемые напряжением натриевые каналы в той части аксона, которая находится ближе всего к телу клетки, активируются благодаря недавно деполяризованному телу клетки. Это позволяет положительно заряженным ионам натрия течь в отрицательно заряженный аксон и деполяризировать окружающий аксон. Мы можем думать о том, что каналы открываются как домино, падающие вниз — как только один канал открывается и пропускает положительные ионы, он создает основу для того, чтобы каналы вниз по аксону делали то же самое. Хотя эта стадия известна как деполяризация, нейрон фактически переходит равновесие и становится положительно заряженным, когда потенциал действия проходит!
Реполяризация — возвращает клетку к потенциалу покоя. Ворота инактивации натриевых каналов закрываются, останавливая приток положительных ионов внутрь. В то же время калиевые каналы открываются. В клетке гораздо больше калия, чем наружу, поэтому, когда эти каналы открываются, выходит больше калия, чем поступает. Это означает, что клетка теряет положительно заряженные ионы и возвращается обратно в состояние покоя.
Гиперполяризация — делает клетку более негативной, чем ее типичный мембранный потенциал в покое. Когда потенциал действия проходит, калиевые каналы остаются открытыми немного дольше и продолжают пропускать положительные ионы из нейрона. Это означает, что ячейка временно гиперполяризована или становится еще более отрицательной, чем ее состояние покоя. Когда калиевые каналы закрываются, натриево-калиевый насос восстанавливает состояние покоя.
Потенциал действия в различных типах клеток
Потенциал действия в мышечных тканях
Потенциал действия в скелетных мышечных клетках аналогичный потенциала действия в нейронах. Потенциал покоя в них как правило -90мВ, что меньше, чем потенциал покоя типовых нейронов. Потенциал действия мышечных клеток длится примерно 2-4 мс, абсолютный рефрактерный период составляет примерно 1-3 мс, а скорость проводимости вдоль мышц примерно 5 м / с.
Потенциал действия в сердечных тканях
Потенциал действия клеток рабочего миокарда состоит из фазы быстрого деполяризации, начальной быстрой реполяризации, которая переходит в фазу медленной реполяризации (фаза плато), и фазы быстрой конечной реполяризации. Фаза быстрой деполяризации обусловлена резким повышением проницаемости мембраны для ионов натрия, вызывает быстрый входящий натриевый ток, при достижении мембранного потенциала 30-40 мВ инактивируется и в дальнейшем главную роль играют кальциевый ионный ток. Деполяризация мембраны вызывает активацию кальциевых каналов, в результате чего возникает дополнительный Деполяризующий входящий кальциевый ток.
Потенциал действия в сердечной ткани играет важную роль в координации сокращений сердца.