Атф-синтаза
Содержание:
Превращение веществ в клетке
Пищевые вещества, попавшие в организм, вовлекаются в процессы метаболизма.
Метаболизм
Метаболизм представляет собой совокупность взаимосвязанных и сбалансированных процессов, включающих разнообразные химические превращения в организме.
Синтез веществ в клетке
Анаболизм
Реакции синтеза, осуществляющиеся с потреблением энергии, составляют основу анаболизма (пластического обмена, или ассимиляции).
Процесс синтеза веществ = пластический обмен = ассимиляция = анаболизм
Чтобы что-то построить, надо затратить энергию — этот процесс идет с поглощением энергии.
| Тип организма | Пример пластического обмена |
| Автотрофы | Фотосинтез |
| Гетеротрофы | Синтез белка |
Катаболизм
Противоположный анаболизму процесс –
Катаболизм
катаболизм – процесс расщепления веществ с высвобождением энергии (энергетический обмен, или диссимиляция).
Процесс расщепления = энергетический обмен = диссимиляция = катаболизм
Это процесс, при котором сложные вещества разлагаются на простые. “Ломать — не строить”, так что энергия при этом выделяется.
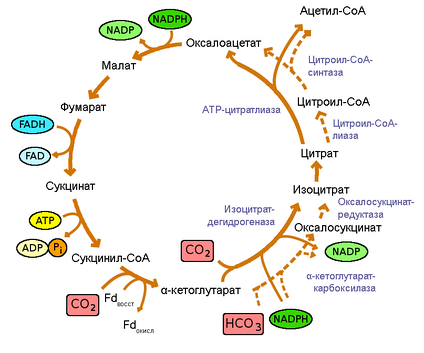
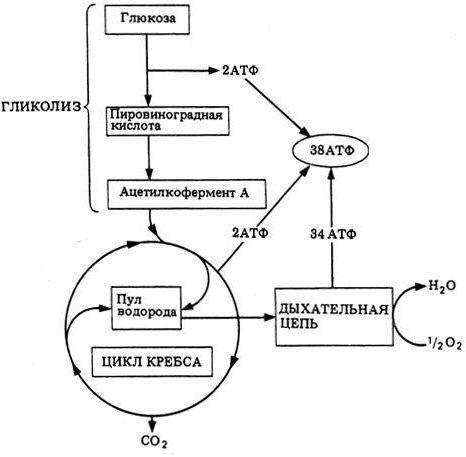
В основном, это реакции окисления, происходят они в митохондриях, самый простой пример — дыхание. При дыхании сложные органические вещества расщепляются до простых, выделяется углекислый газ и энергия.
Вообще, эти два процесса взаимосвязаны и переходят один в другой. Суммарно уравнение метаболизма — обмена веществ в клетке — можно записать так:
катаболизм + анаболизм = обмен веществ в клетке = метаболизм
Рассмотрим эти процессы подробнее.
Энергетический обмен = Диссимиляция = Катаболизм
Этот процесс идет в несколько этапов, и нам нужно рассмотреть, как он проходит в различных организмах.
Организмов будет всего 2 — многоклеточный (человек, например) и одноклеточный (растительный и животный).
И запомните, сочетание букв АТФ (аденозинтрифосфат) — означает “энергию”. Просто эта энергия заключена в молекуле.
Анаболизм
Катаболические пути влияют на превращение пищевых материалов в необратимые интермедиаты. Анаболические пути, с другой стороны, представляют собой последовательности катализируемых ферментами реакций, в которых компонентные строительные блоки больших молекул или макромолекул (например, белков, углеводов и жиров) образуются из одних и тех же промежуточных продуктов.
Таким образом, катаболические маршруты имеют четко определенные начала, но не имеют однозначно идентифицируемых конечных продуктов; анаболические пути, с другой стороны, приводят к четко различимым конечным продуктам от диффузных начал. Два типа пути связаны между собой реакциями переноса фосфатов, в которых участвуют АДФ (аденозиндифосфат), АМФ (аденонмонофосфорная кислота) и АТФ, а также посредством переносов электронов, которые позволяют уменьшить количество восстанавливающих эквивалентов (то есть атомов водорода или электронов), которые высвобождаются во время катаболических реакций. используется для биосинтеза.
Но, хотя катаболические и анаболические пути тесно связаны между собой, и общий эффект одного типа маршрута явно противоположен другому, у них мало общих шагов.
Анаболический путь для синтеза конкретной молекулы обычно начинается с промежуточных соединений, весьма отличных от тех, которые образуются в результате катаболизма этой молекулы; например, микроорганизмы катаболизируют ароматические (то есть содержащие кольцевую или циклическую структуру) аминокислоты до ацетилкофермента А и промежуточного соединения цикла Кребса. Биосинтез этих аминокислот, однако, начинается с соединения, полученного из пирувата, и промежуточного соединения метаболизма пентозы (общее название для сахаров с пятью атомами углерода). Аналогично, гистидин синтезируется из пентозного сахара, но катаболизируется в α-оксоглутарат.
Даже в тех случаях, когда продукт катаболизма используется в анаболическом пути, возникают различия; таким образом, жирные кислоты, которые катаболизируются в ацетил-кофермент А, синтезируются не непосредственно из ацетил-кофермента А, а из его производного, малонил-кофермента А ( см. ниже липидные компоненты ). Кроме того, даже ферменты, которые катализируют очевидно идентичные стадии в катаболических и анаболических путях, могут проявлять различные свойства. В общем, следовательно, путь вниз (катаболизм) отличается от пути вверх ( анаболизм ). Эти различия важны, потому что они позволяют регулировать катаболические и анаболические процессы в клетке.
Как увеличить выработку АТФ
Количество молекул «энергетической» кислоты зависит от количества митохондрий в организме. Есть несколько способов повысить число митохондрий:
- Физические нагрузки. Регулярные тренировки истощают запасы энергии, поэтому требуют большое количество митохондрий для производства АТФ.
- Холод. Опыты на крысах показали, что низкие температуры влияют на число митохондрий.
- Диета с низким содержанием углеводов и высоким содержанием жиров заставляет организм использовать для выработки энергии жиры.
- Добавки, которые помогают митохондриям вырабатывать энергию более эффективно. К ним относят, например, Коэнзим Q10.
Безусловно, организм сам знает, когда выделять нужные ему вещества. Но если вы регулярно занимаетесь спортом или желаете отсрочить наступление старости, то вам стоит ему немного помочь.
АТФ-синтаза у разных организмов
Расположение АТФ-синтазы в митохондрии
АТФ-синтаза растений
У растений АТФ-синтаза CF1FO присутствует в хлоропластах. Она встроена в мембрану тилакоида, причём компонент CF1 выступает в строму, где протекают темновые реакции фотосинтеза (также называемые светонезависимыми реакциями цикла Кальвина). Структура и механизм катализа АТФ-синтазы хлоропластов почти такая же, как и в митохондриях. Однако электрохимический потенциал у хлоропластов формируется не дыхательной электрон-транспортной цепочкой, а другими комплексами — фотосистемой II и цитохромным комплексом b6/f.
Механизм изменения участка связывания
Основные субъединицы (α3β3) компонента F1 соединены дополнительной боковой ножкой с неподвижным участком FO, что предотвращает их вращение вместе с γ-субъединицей.
Структура неповрежденной АТФ-синтазы с низкой точностью выявлена при помощи электронной криомикроскопии (ЭКМ). Показано, что боковая ножка — это гибкая перемычка, похожая на канат, наматывающаяся на комплекс во время его работы.
В определенных условиях каталитическая реакция может протекать в обратном направлении, при этом гидролиз АТФ вызывает прокачку протонов через мембрану.
В механизме изменения участка связывания задействован активный участок β-субъединицы, последовательно проходящий через три состояния.
В «открытом» состоянии АДФ и фосфат подходят к активному участку. Затем белок охватывает эти молекулы и свободно связывается с ними («свободное» состояние). Следующее изменение формы белка прижимает молекулы друг к другу («тесное» состояние), что приводит к формированию АТФ. Наконец, активный участок снова переходит в «открытое» состояние, освобождает АТФ и связывает следующую молекулу АДФ и фосфата, после чего цикл производства АТФ повторяется.
Структура и номенклатура
Имеющаяся в митохондриях АТФ-синтаза F1FO очень хорошо исследована.
- компонент FO — трасмембранный домен,
- компонент F1 находится вне мембраны, в матриксе.
АТФ-синтазный комплекс FOF1 по форме напоминает плодовое тело гриба, у которого компонент F1 — это шляпка, ножка — это γ-субъединица компонента F1, а «корни» гриба — компонент FO, заякоренный в мембране.
В структурно-функциональном плане АТФ-синтетаза состоит из двух крупных фрагментов, обозначаемых символами F1 и FO. Первый из них (фактор сопряжения F1) обращён в сторону матрикса митохондрии и заметно выступает из мембраны в виде сферического образования высотой 8 нм и шириной 10 нм. Он состоит из девяти субъединиц, представленных пятью типами белков. Полипептидные цепи трёх субъединиц α и стольких же субъединиц β уложены в похожие по строению белковые глобулы, которые вместе образуют гексамер (αβ)3, имеющий вид слегка приплюснутого шара. Подобно плотно уложенным долькам апельсина, последовательно расположенные субъединицы α и β образуют структуру, характеризующуюся осью симметрии третьего порядка с углом поворота 120°. В центре этого гексамера находится субъединица γ, которая образована двумя протяжёнными полипептидными цепями и напоминает слегка деформированный изогнутый стержень длиной около 9 нм. При этом нижняя часть субъединицы γ выступает из шара на 3 нм в сторону мембранного комплекса F. Также внутри гексамера находится минорная субъединица ε, связанная с γ. Последняя (девятая) субъединица обозначается символом δ и расположена на внешней стороне F1.
Мембранная часть АТФ-синтетазы, называемая фактором сопряжения FO, представляет собой гидрофобный белковый комплекс, пронизывающий мембрану насквозь и имеющий внутри себя два полуканала для прохождения протонов водорода (ядер протия). Всего в состав комплекса FO входит одна белковая субъединица типа а, две копии субъединицы b, а также от 9 до 12 копий мелкой субъединицы c. Субъединица а (молекулярная масса 20 кДа) полностью погружена в мембрану, где образует шесть пересекающих её α-спиральных участков. Субъединица b (молекулярная масса 30 кДа) содержит лишь один сравнительно короткий погружённый в мембрану α-спиральный участок, а остальная её часть заметно выступает из мембраны в сторону F1 и закрепляется за расположенную на её поверхности субъединицу δ. Каждая из 9-12 копий субъединицы c (молекулярная масса 6-11 кДа) представляет собой сравнительно небольшой белок из двух гидрофобных α-спиралей, соединённых друг с другом короткой гидрофильной петлёй, ориентированной в сторону F1, а все вместе образуют единый ансамбль, имеющий форму погружённого в мембрану цилиндра. Выступающая из комплекса F1 в сторону FO субъединица γ как раз и погружена внутрь этого цилиндра и достаточно прочно зацеплена за него.
Номенклатура фермента имеет традиционное происхождение, поэтому довольно непоследовательна.
Обозначение компонента F1 является сокращением от «Fraction 1» (часть 1), а символом FO (в индексе записана буква O, а не ноль) обозначался участок связывания олигомицина.
Некоторые субъединицы фермента имеют также буквенные обозначения:
- Греческие: α, β, γ, δ, ε
- Латинские: a, b, c, d, e, f, g, h
Другие — более сложные обозначения:
- F6 (от «Fraction 6»)
- OSCP — белок, чувствительный к олигомицину (от англ. the oligomycin sensitivity conferral protein),
- A6L (названный так по названию гена, кодирующего его в митохондриальном геноме)
- IF1 (фактор ингибирования 1),
Компонент F1 достаточно велик (диаметр его составляет 9 нм), чтобы быть видимым в трансмиссионный электронный микроскоп при негативном окрашивании.
Частичками F1 усеяна внутренняя митохондриальная мембрана. Изначально считалось, что они содержат весь дыхательный аппарат митохондрии. Однако после долгих экспериментов группа Эфраима Рекера (впервые выделившая компонент F1 в 1961) показала, что эти частички связаны с АТФазной активностью в том числе и в разделённых митохондриях, и в субмитохондриальных частицах, формирующихся при ультразвуковом воздействии на митохондрии. Множество дальнейших исследований в разных лабораториях подтвердили эту АТФазную активность.
АТФ-синтаза у разных организмов[править | править код]
Расположение АТФ-синтазы в митохондрии
АТФ-синтаза растенийправить | править код
У растений АТФ-синтаза CF1FO присутствует в хлоропластах. Она встроена в мембрану тилакоида, причём компонент CF1 выступает в строму, где протекают темновые реакции фотосинтеза (также называемые светонезависимыми реакциями цикла Кальвина). Структура и механизм катализа АТФ-синтазы хлоропластов почти такая же, как и в митохондриях. Однако электрохимический потенциал у хлоропластов формируется не дыхательной электрон-транспортной цепочкой, а другими комплексами — фотосистемой II и цитохромным комплексом b6/f.
Структура и формула АТФ
Если говорить об АТФ более подробно
, то это молекула, которая даёт энергию всем процессам, происходящим в организме, в том числе она же даёт энергию для движения. При расщеплении молекулы АТФ происходит сокращение мышечного волокна, вследствие чего выделяется энергия, позволяющая произойти сокращению. Синтезируется Аденозинтрифосфат из инозина — в живом организме.
Для того чтобы дать организму энергию Аденозинтрифосфату необходимо пройти несколько этапов. Вначале отделяется один из фосфатов — с помощью специального коэнзима. Каждый из фосфатов даёт десять калорий. В процессе вырабатывается энергия и получается АДФ (аденозин дифосфат).
Если организму для действия нужно больше энергии
, то отделяется ещё один фосфат. Тогда формируется АМФ (аденозин монофосфат). Главный источник для выработки Аденозинтрифосфата — это глюкоза, в клетке она расщепляется на пируват и цитозол. Аденозинтрифосфат насыщает энергией длинные волокна, которые содержат протеин — миозин. Именно он формирует мышечные клетки.
В моменты, когда организм отдыхает, цепочка идёт в обратную сторону, т. е. формируется Аденозин Три-Фосфорная кислота. Опять же в этих целях используется глюкоза. Созданные молекулы Аденозинтрифосфата будут вновь использоваться, как только это станет необходимо. Когда энергия не нужна, она сохраняется в организме и высвобождается как только это потребуется.
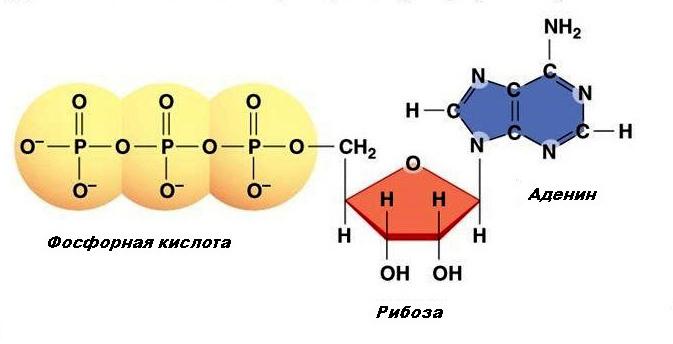
Молекула АТФ состоит из нескольких, а точнее, трёх компонентов:
- Рибоза — это пятиуглеродный сахар, такой же лежит в основе ДНК.
- Аденин — это объединённые атомы азота и углерода.
- Трифосфат.
В самом центре молекулы Аденозинтрифосфата находится молекула рибозы, а её край является основной для аденозина. С другой стороны рибозы расположена цепочка из трёх фосфатов.
Эволюция АТФ-синтазы
Эволюция АТФ-синтазы считается примером модульной эволюции, при которой две субъединицы, каждая обладающая своими функциями, соединились и получили новые функции.
Гексамер α3β3, входящий в состав компонента F1 проявляет существенное сходство с гексамерной ДНК-геликазой. Оба типа ферментов образуют кольцо с вращательной симметрией 3 порядка, обладающее центральной пóрой. Действие каждого из них также зависит от относительного вращения макромолекулы внутри поры: геликазы используют спиральную форму ДНК для движения вдоль неё и для обнаружения суперскручивания, тогда как α3β3-гексамер использует изменения своей конфигурации из-за вращения γ-субъединицы для осуществления каталитической реакции.
Протонный мотор компонента FO проявляет большое функциональное сходство с протонными моторами жгутиков. И там, и там присутствует кольцо из множества небольших богатых α-спиралями белков, вращающихся относительно соседних неподвижных белков за счёт энергии протонного градиента. Это, конечно, очень зыбкое сходство, так как структура жгутиковых моторов гораздо сложнее, чем FO, а вращающееся белковое кольцо гораздо крупнее и состоит из 30 субъединиц против 10, 11 или 14, входящих в состав компонента FO.
Теория молекулярной эволюции предполагает, что две субъединицы с независимыми функциями — ДНК-геликаза с дополнительным АТФ-азным действием и протонный мотор — смогли соединяться, причём вращение мотора вызывало проявление АТФ-азной активности геликазы. Или же, наоборот, в первичной связке ДНК-геликазы и протонного мотора гидролиз АТФ на геликазе заставлял работать протонный мотор. Это соединение затем постепенно оптимизировалось, получило возможность катализировать обратную реакцию и через какое-то время превратилось в сложную АТФ-синтазу, существующую в настоящее время. Однако, до сих пор неясен механизм происхождения протонного мотора, который без геликазы или других комплексов не представляет никакой пользы.
Понятие метаболизма
Метаболизм — совокупность всех химических реакций, протекающих в живом организме. Значение метаболизма состоит в создании необходимых организму веществ и обеспечении его энергией.
Выделяют две составные части метаболизма — катаболизм и анаболизм.
Составные части метаболизма
Часть
Характеристика
Примеры
Затраты энергии
Катаболизм (энергетический обмен, диссимиляция)
Совокупность химических реакций, приводящих к образованию простых веществ из более сложных
Гидролиз полимеров до мономеров и расщепление последних до низкомолекулярных соединений углекислого газа, воды, аммиака и других веществ
Энергия выделяется
Анаболизм (пластический обмен, ассимиляция)
Совокупность химических реакций синтеза сложных веществ из более простых
Образование углеводов из углекислого газа и воды в процессе фотосинтеза, реакции матричного синтеза
Энергия поглощается
Процессы пластического и энергетического обмена неразрывно связаны между собой. Все синтетические (анаболические) процессы нуждаются в энергии, поставляемой в ходе реакций диссимиляции. Сами же реакции расщепления (катаболизма) протекают лишь при участии ферментов, синтезируемых в процессе ассимиляции.
Роль ФТФ в метаболизме
Энергия, высвобождающаяся при распаде органических веществ, не сразу используется клеткой, а запасается в форме высокоэнергетических соединений, как правило, в форме аденозинтрифосфата (АТФ). По своей химической природе АТФ относится к мононуклеотидам.
АТФ (аденозинтрифосфорная кислота) — мононуклеотид, состоящий из аденина, рибозы и трёх остатков фосфорной кислоты, соединяющихся между собой макроэргическими связями.
В этих связях запасена энергия, которая высвобождается при их разрыве:
АТФ + H2O → АДФ + H3PO4 + Q1
АДФ + H2O → АМФ + H3PO4 + Q2
АМФ + H2O → аденин + рибоза + H3PO4 + Q3,
где АТФ — аденозинтрифосфорная кислота; АДФ — аденозиндифосфорная кислота; АМФ — аденозинмонофосфорная кислота; Q1 = Q2 = 30,6 кДж; Q3 = 13,8 кДж.
Запас АТФ в клетке ограничен и пополняется благодаря процессу фосфорилирования. Фосфорилирование — присоединение остатка фосфорной кислоты к АДФ (АДФ + Ф → АТФ). Он происходит с разной интенсивностью при дыхании, брожении и фотосинтезе. АТФ обновляется чрезвычайно быстро (у человека продолжительность жизни одной молекулы АТФ менее 1 мин).
Энергия, накопленная в молекулах АТФ, используется организмом в анаболических реакциях (реакциях биосинтеза). Молекула АТФ является универсальным хранителем и переносчиком энергии для всех живых существ.
Световая фаза фотосинтеза
В световой фазе фотосинтеза происходит синтез АТФ и НАДФ·H2 за счет лучистой энергии. Это происходит на тилакоидах хлоропластов, где пигменты и ферменты образуют сложные комплексы для функционирования электрохимических цепей, по которым передаются электроны и отчасти протоны водорода.
Электроны в конечном итоге оказываются у кофермента НАДФ, который, заряжаясь отрицательно, притягивает к себе часть протонов и превращается в НАДФ·H2. Также накопление протонов по одну сторону тилакоидной мембраны и электронов по другую создает электрохимический градиент, потенциал которого используется ферментом АТФ-синтетазой для синтеза АТФ из АДФ и фосфорной кислоты.
Главными пигментами фотосинтеза являются различные хлорофиллы. Их молекулы улавливают излучение определенных, отчасти разных спектров света. При этом некоторые электроны молекул хлорофилла переходят на более высокий энергетический уровень. Это неустойчивое состояние, и по-идее электроны путем того же излучения должны отдать в пространство полученную из вне энергию и вернуться на прежний уровень. Однако в фотосинтезирующих клетках возбужденные электроны захватываются акцепторами и с постепенным уменьшением своей энергии передаются по цепи переносчиков.
На мембранах тилакоидов существуют два типа фотосистем, испускающих электроны при действия света. Фотосистемы представляют собой сложный комплекс большей частью хлорофильных пигментов с реакционным центром, от которого и отрываются электроны. В фотосистеме солнечный свет ловит множество молекул, но вся энергия собирается в реакционном центре.
Электроны фотосистемы I, пройдя по цепи переносчиков, восстанавливают НАДФ.
Энергия электронов, оторвавшихся от фотосистемы II, используется для синтеза АТФ. А сами электроны фотосистемы II заполняют электронные дырки фотосистемы I.
Дырки второй фотосистемы заполняются электронами, образующимися в результате фотолиза воды. Фотолиз также происходит при участии света и заключается в разложении H2O на протоны, электроны и кислород. Именно в результате фотолиза воды образуется свободный кислород. Протоны участвуют в создании электрохимического градиента и восстановлении НАДФ. Электроны получает хлорофилл фотосистемы II.
Примерное суммарное уравнение световой фазы фотосинтеза:
H2O + НАДФ + 2АДФ + 2Ф → ½O2 + НАДФ · H2 + 2АТФ
Циклический транспорт электронов
Выше описана так называемый нецикличная световая фаза фотосинтеза. Есть еще циклический транспорт электронов, когда восстановления НАДФ не происходит. При этом электроны от фотосистемы I уходят на цепь переносчиков, где идет синтез АТФ. То есть эта электрон-транспортная цепь получает электроны из фотосистемы I, а не II. Первая фотосистема как бы реализует цикл: в нее возвращаются ей же испускаемые электроны. По дороге они тратят часть своей энергии на синтез АТФ.
Фотофосфорилирование и окислительное фосфорилирование
Световую фазу фотосинтеза можно сравнить с этапом клеточного дыхания — окислительным фосфорилированием, которое протекает на кристах митохондрий. Там тоже происходит синтез АТФ за счет передачи электронов и протонов по цепи переносчиков. Однако в случае фотосинтеза энергия запасается в АТФ не для нужд клетки, а в основном для потребностей темновой фазы фотосинтеза. И если при дыхании первоначальным источником энергии служат органические вещества, то при фотосинтезе – солнечный свет. Синтез АТФ при фотосинтезе называется фотофосфорилированием, а не окислительным фосфорилированием.