Цикл кребса простым языком
Содержание:
- Разница между гликолизом и глюконеогенезом
- Занятие № 12. «Цикл трикарбоновых кислот»
- Дыхательная цепь
- Описание процесса
- История изучения
- Гликолиз
- Этапы энергетического обмена
- Значение и функции
- Строение ферментативных комплексов дыхательной цепи
- Метаболизм
- Метаболизм веществ
- Регуляция цикла Кребса[править | править код]
- Клеточное дыхание
- Химизм
- Ферменты
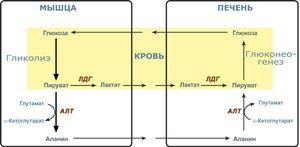
Разница между гликолизом и глюконеогенезом
Определение
Гликолиз: Набор реакций, которые превращают глюкозу в две молекулы пирувата, известен как гликолиз.
Глюконеогенез: Глюконеогенез — это производство неуглеводных источников глюкозы, таких как глицерин, аминокислоты и лактат.
Сырье
Гликолиз: Сырьем для гликолиза является глюкоза.
Глюконеогенез: Сырьем для глюконеогенеза является лактат, аминокислоты, такие как аланин и глицерин.
Вхождение
Гликолиз: Гликолиз происходит в цитоплазме всех клеток.
Глюконеогенез: Глюконеогенез происходит как в митохондриях, так и в цитоплазме.
В тканях
Гликолиз: Гликолиз происходит практически во всех клетках организма.
Глюконеогенез: Глюконеогенез происходит в печени и почках.
метаболизм
Гликолиз: Гликолиз — это катаболический процесс, когда молекулы глюкозы распадаются на две молекулы пирувата.
Глюконеогенез: Глюконеогенез — это анаболический процесс, при котором две молекулы пирувата соединяются вместе, образуя молекулу глюкозы.
Использование энергии
Гликолиз: Гликолиз является экзергонической реакцией, при которой образуются два АТФ.
Глюконеогенез: Глюконеогенез является эндергонической реакцией, где шесть АТФ используются на одну молекулу глюкозы.
переписка
Гликолиз: Гликолиз происходит через десять реакций.
Глюконеогенез: Две по существу необратимые реакции в гликолитическом пути обойдены четырьмя новыми реакциями в глюконеогенезе.
Шаг ограничения скорости
Гликолиз: Ферменты, участвующие в стадиях ограничения скорости, представляют собой гексокиназу, фосфофруктокиназу и пируваткиназу.
Глюконеогенез: Ферменты, участвующие в стадиях ограничения скорости, представляют собой пируваткарбоксилазу, фосфоенолпируваткарбоксикиназу, фруктозу 1,2-бисфосфатазу, глюкозо-6-фосфатфосфатазу.
Заключение
Гликолиз и глюконеогенез являются двумя процессами, участвующими в метаболизме глюкозы. Глюкоза является источником энергии практически всех форм жизни на Земле. Глюкоза расщепляется для выработки энергии в форме АТФ во время процесса, называемого клеточным дыханием. Гликолиз является первым этапом клеточного дыхания, расщепляющим шесть углеродных глюкоз на две молекулы пирувата, каждая из которых содержит три атома углерода. Гликолиз происходит в цитоплазме практически всех клеток организма. Во время голодания уровни глюкозы в крови снижаются, и печень и почки начинают вырабатывать глюкозу из неуглеводных производных, таких как аминокислоты, глицерин и лактат, в процессе, называемом глюконеогенезом. Глюконеогенез и гликолиз являются взаимно регулируемыми событиями, поддерживая постоянный уровень глюкозы в крови. Основное различие между гликолизом и глюконеогенезом заключается в их типах обмена веществ в организме.
Ссылка:1. Берг, Джереми М. «Гликолизис — путь преобразования энергии во многих организмах». Биохимия. 5-е издание. Национальная медицинская библиотека США, 1 января 1970 г. Веб. 06 апреля 2017 г. 2. Берг, Джереми М. «Резюме». Биохимия. 5-е издание, Национальная медицинская библиотека США, 1 января 1970 г. Веб. 06 апреля 2017 г.
Занятие № 12. «Цикл трикарбоновых кислот»
Цель занятия: изучить механизм некоторых реакций цикла Кребса. Освоить метод количественного определения пировиноградной кислоты в моче.
ВОПРОСЫ К КОНТРОЛЬНОЙ РАБОТЕ:
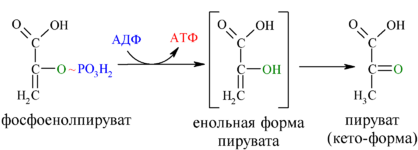
1. Окислительное декарбоксилирование пирувата как предварительный этап цикла лимонной кислоты. Перечислите витамины и коферменты, задействованные в этом процессе.
2. Реакции цикла лимонной кислоты. Что определяет общее направление реакций в цикле? В какой части клетки протекает этот процесс? Почему?
3. Какие коферменты и витамины участвуют в цикле Кребса? Объясните, как они работают, с указанием конкретных реакций.
4. Расскажите о реакциях цикла Кребса, в результате которых образуются НAДH2 и ФAДН2. Какова дальнейшая судьба этих соединений?
5. Функции цикла трикарбоновых кислот. Объясните, какое значение для цикла лимонной кислоты имеет анаплеротическая реакция?
6. Энергетический выход цикла трикарбоновых кислот. Сколько молекул ATФ образуется в ходе оборота через цикл одной молекулы лимонной кислоты? Все ли молекулы ATФ, образующиеся при полном окислении активного ацетила, синтезируются путем окислительного фосфорилирования. Как регулируется скорость цикла?
Дата выполнения ________ Балл ____ Подпись преподавателя ____________
Экспериментальная работа.
Одним из методов количественного анализа в биохимии является фотокалориметрия. Метод основан на измерении оптической плотности окрашенных растворов, которые получают при взаимодействии субстрата с различными химическими агентами. Концентрация субстрата пропорциональна степени окрашивания раствора.
Перед началом лабораторных опытов ознакомьтесь с устройством ФЭК и правилами работы на нем.
Опыт 1. Определение концентрации пировиноградной кислоты (ПВК) в моче.
Приборы. 1. Штатив с пробирками.
2. Набор пипеток.
3. Фотоколориметр.
4. Кюветы, 0,5 см.
Реактивы. 1. Дистиллированная вода.
2. Моча.
3. Гидроксид натрия, 10%-ный раствор.
4. 2,4-динитрофенилгидразин, раствор.
ПВК крови конденсируется с 2,4-динитрофинилгидразином с образованием гидразона, который в щелочной среде дает коричнево-красный цвет раствора. По интенсивности его окраски судят о содержании ПВК.
1. В три пробирки внесите реактивы согласно следующей таблице:
| Раствор | Опытная проба, мл | Контрольная проба, мл |
| Исследуемый образец | — | |
| Калибр. раствор пирувата* | — | — |
| Дистиллированная вода | — | |
| 2,4-динитрофенилгидразин | 0,8 | 0,8 |
2. Содержимое пробирок на 15 мин поместите в темное место при комнатной температуре.
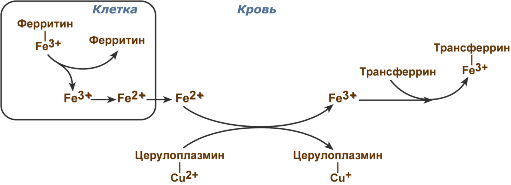
Дыхательная цепь
Организация дыхательной цепи
Протекает на внутренних мембранах митохондрий, где расположен ряд ферментов в определенной последовательности (дыхательная цепь). Атомы водорода попадают на мембраны митохондрий. Через ряд этапов происходит с их помощью восстановление АТФ.
Высвобожденный в цикле Кребса водород объединяется с НАД (никотинамидадениндинуклеотидом). Образуется восстановленная форма НАД • Н. Далее НАД • Н окисляется до НАД+ , Н+ и электрона (е) и транспортируется на внутреннюю поверхность мембраны митохондрий. Ионы водорода накапливаются на внешней поверхности внутренней мембраны, а электроны с помощью переносчиков попадают на внутреннюю поверхность внутренней мембраны. На внутренней поверхности уменьшается количество ионов водорода, образуется вода:
4Н+ + 4 + 02 → 2Н20.
Возникает разница электрических потенциалов, концентраций ионов водорода по разные стороны внутренней мембраны.
АДФ и фосфорная кислота восстанавливают АТФ с помощью особой ферментной системы, которая использует для этого разницу электрических потенциалов, различие концентраций ионов водорода. Эта ферментная система переводит ионы водорода на внутреннюю поверхность внутренней мембраны с внешней поверхности. Процесс образования АТФ из АДФ и фосфорной кислоты называется окислительным фосфорилированием. Процесс перенесения электрона по дыхательной цепи митохондрий имеет название сопряжение окисления.
При окислении двух молекул молочной кислоты выделяется энергия, которая обеспечивает образование 36 молекул АТФ:
2С3Н603 + 602 + 36АДФ + 36Н3Р04 → 6С02 + 36АТФ + 36Н20.
Суммарное уравнение энергетического обмена:
С6Н1206 + 602 + 38АДФ + 38Н3Р04 → 6С02 + 38АТФ + 44Н20.
Выделяется почти 2,8 тыс. кДж энергии. 1596 кДж (55 %) – запасается в виде макроэргических связей АТФ. Оставшиеся (45 %) рассеиваются в виде тепла.
Описание процесса
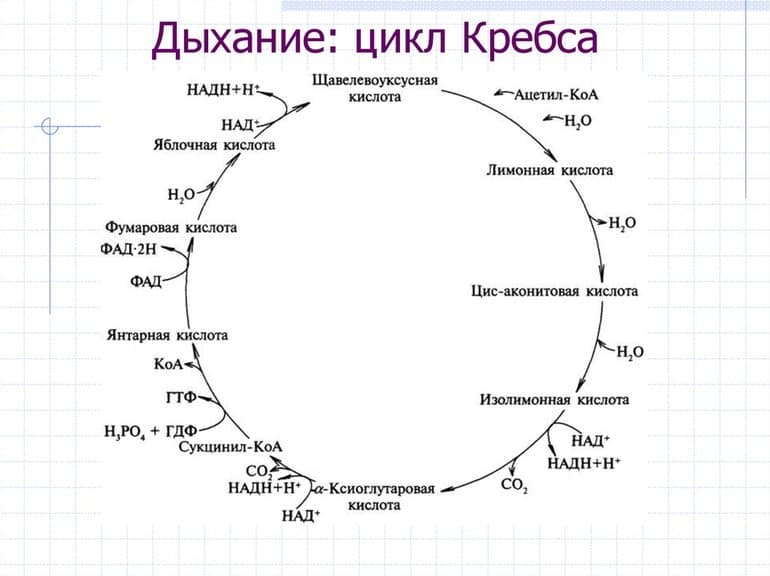
Протекание ЦК достаточно экономно с точки зрения энергозатрат. Такой эффект достигается благодаря тому, что он связывает два метаболических направления. В процесс вовлекаются вещества, подлежащие утилизации, которые либо служат энергетическим «топливом», либо возвращаются в круг анаболизма. Подготовительная стадия ЦК заключается в распаде глюкозы, аминокислот и жирных кислот на молекулы пирувата или лактата.
Органеллы митохондрий способны преобразовывать пируват в ацетильный остаток (ацетил-коэнзим А или ацетил-КоА), представляющий собой вместе с тиольной группой, которая может его переносить, кофермент А. Некоторое соединения могут сразу распадаться до ацетил-КоА, минуя стадию пирувата. При этом пировиноградная кислота может вовлекаться непосредственно в ЦК, не преобразуясь в ацетил-КоА.
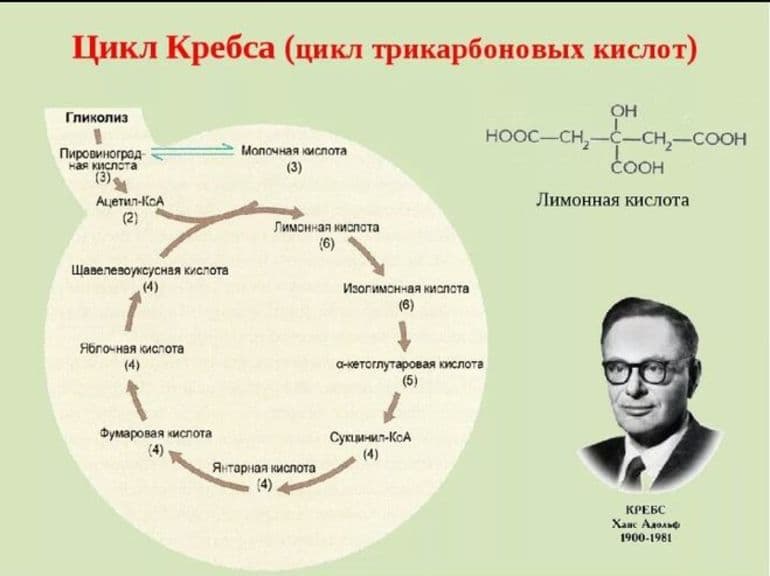
Начальные этапы
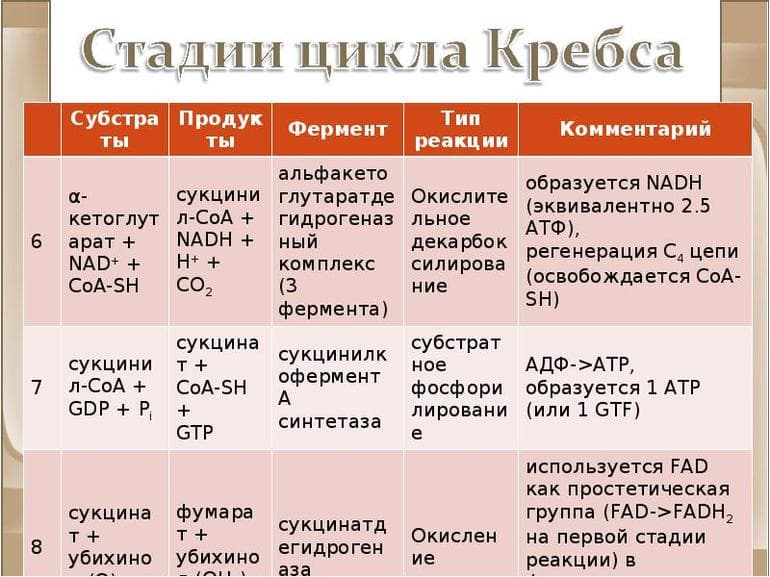
Первая стадия необратима и состоит из конденсации ацетил-КоА с четырехуглеродным веществом — оксалоацетатом (щавелевоуксусной кислотой или ЩУК), что приводит к образованию шестиуглеродного цитрата (лимонной кислоты). Во время реакции метильная группа ацетил-КоА соединяется с карбонильной группой ЩУК. Благодаря быстрому гидролизу промежуточного соединения цитроил-КоА этот этап проходит без затрат энергии извне.
На второй стадии образуется изоцитрат (изолимонная кислота) из цитрата через цис-аконитат. Это реакция обратимой изомеризации через образование промежуточной трикарбоновой кислоты, в которой катализатором выступает фермент аконитатгидратаза.
Далее происходит дегидрирование и декарбоксилирование изоцитрата до промежуточного соединения оксалосукцинат с выделением углекислого газа. После декарбоксилирования оксалосукцината образуется енольное соединение, которое перестраивается и превращается в пятиуглеродную кислоту — α-кетоглутарат (оксоглутарата), чем и завершает третью ступень ЦК. Четвертый этап — α-кетоглутарат декарбоксилирует и реагирует с ацетил-КоА. При этом получается сукцинил-КоА, соединение янтарной кислоты и коэнзима-А, выделяется СО2.
Замыкание цикла
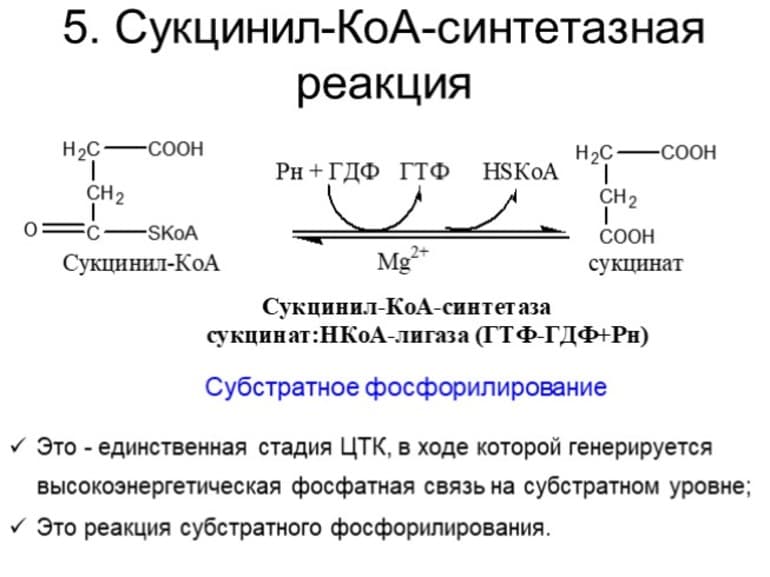
На пятой стадии сукцинил-КоА преобразуется в сукцинат (янтарную кислоту). Для этого этапа характерно субстратное фосфолирование, подобное синтезу АТФ при гликолизе. Введение в ЦК фосфорной группы РО3 становится возможным благодаря присутствию фермента ГДФ (гуанозиндифосфата) или АДФ (аденозиндифосфата), которые в процессе синтеза сукцината из дифосфатов становятся трифосфатами.
Начиная с шестой стадии, цикл начинает постепенно замыкаться. Сначала сукцинат под действием каталитического фермента сукцинатдегидрогеназы дегидрирует до фумарата. Дальнейшее дигидрирование приводит к седьмому этапу — образованию L-малата (яблочной кислоты) из фуратата через переходное соединение с карбанионом.
История изучения
Биологическая роль некоторых реакций цикла Кребса (ЦК) была изучена американским биохимиком венгерского происхождения Альбертом Сент-Дьердьи. В частности, он выделил ключевой компонент ЦТК — фумарат. Исследования в этом направлении продолжил Ганс Кребс. В итоге он установил всю последовательность реакций и соединений, образующиеся на всех этапах процесса. Ученый не смог определить, с преобразования какой кислоты начинается цикл — лимонной или изолимонной. Сейчас известно, что это лимонная кислота. Поэтому ЦК называют также цитратным или циклом лимонной кислоты.
Гликолиз
Анаэробное превращение глюкозы локализуется в цитозоле и включает два этапа из 11 ферментативных реакций.
Первый этап гликолиза
Первый этап гликолиза – подготовительный, здесь происходит затрата энергии АТФ, активация глюкозы и образование из нее триозофосфатов.
Первая реакция гликолиза сводится к превращению глюкозы в реакционно-способное соединение за счет фосфорилирования 6-го, не включенного в кольцо, атома углерода. Эта реакция является первой в любом превращении глюкозы, катализируется гексокиназой.
Вторая реакция необходима для выведения еще одного атома углерода из кольца для его последующего фосфорилирования (фермент глюкозофосфат-изомераза). В результате образуется фруктозо-6-фосфат.
Третья реакция – фермент фосфофруктокиназа фосфорилирует фруктозо-6-фосфат с образованием почти симметричной молекулы фруктозо-1,6-дифосфата. Эта реакция является главной в регуляции скорости гликолиза.
В четвертой реакции фруктозо-1,6-дифосфат разрезается пополам фруктозо-1,6-дифосфат-альдолазой с образованием двух фосфорилированных триоз-изомеров – альдозы глицеральдегида (ГАФ) и кетозы диоксиацетона (ДАФ).
Пятая реакция подготовительного этапа – переход глицеральдегидфосфата и диоксиацетонфосфата друг в друга при участии триозофосфатизомеразы. Равновесие реакции сдвинуто в пользу диоксиацетонфосфата, его доля составляет 97%, доля глицеральдегидфосфата – 3%. Эта реакция, при всей ее простоте, определяет дальнейшую судьбу глюкозы:
- при нехватке энергии в клетке и активации окисления глюкозы диоксиацетонфосфат превращается в глицеральдегидфосфат, который далее окисляется на втором этапе гликолиза,
- при достаточном количестве АТФ, наоборот, глицеральдегидфосфат изомеризуется в диоксиацетонфосфат, и последний отправляется на синтез жиров.
Второй этап гликолиза
Второй этап гликолиза – это освобождение энергии, содержащейся в глицеральдегидфосфате, и запасание ее в форме АТФ.
Шестая реакция гликолиза (фермент глицеральдегидфосфат-дегидрогеназа) – окисление глицеральдегидфосфата и присоединение к нему фосфорной кислоты приводит к образованию макроэргического соединения 1,3-дифосфоглицериновой кислоты и НАДН.
В седьмой реакции (фермент фосфоглицераткиназа) энергия фосфоэфирной связи, заключенная в 1,3-дифосфоглицерате тратится на образование АТФ. Реакция получила дополнительное название – реакция субстратного фосфорилирования, что уточняет источник энергии для получения макроэргической связи в АТФ (от субстрата реакции) в отличие от окислительного фосфорилирования (использование энергии электрохимического градиента ионов водорода на мембране митохондрий).
Восьмая реакция – синтезированный в предыдущей реакции 3-фосфоглицерат под влиянием фосфоглицератмутазы изомеризуется в 2-фосфоглицерат.
Девятая реакция – фермент енолаза отрывает молекулу воды от 2-фосфоглицериновой кислоты и приводит к образованию макроэргической фосфоэфирной связи в составе фосфоенолпирувата.
Десятая реакция гликолиза – еще одна реакция субстратного фосфорилирования – заключается в переносе пируваткиназой макроэргического фосфата с фосфоенолпирувата на АДФ и образовании пировиноградной кислоты.
Последняя реакция бескислородного окисления глюкозы, одиннадцатая – образование молочной кислоты из пирувата под действием лактатдегидрогеназы
Важно то, что эта реакция осуществляется только в анаэробных условиях. Эта реакция необходима клетке, так как НАДН, образующийся в 6-й реакции, в отсутствие кислорода не может окисляться в митохондриях
| У плода и детей первых месяцев жизни преобладает анаэробный распад глюкозы, в связи с чем концентрация молочной кислоты в крови у них выше чем у взрослых. |
| При наличии кислорода пировиноградная кислота переходит в митохондрию и превращается в ацетил-S-КоА. |
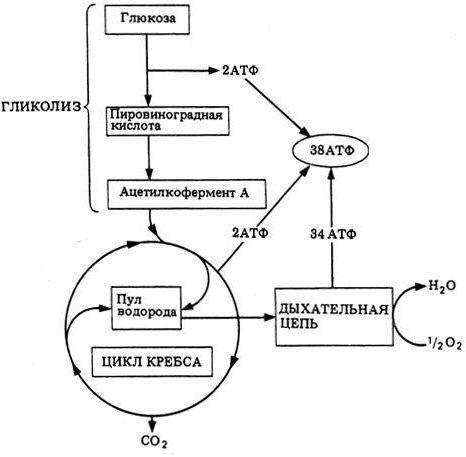
Этапы энергетического обмена
Энергетический обмен состоит из трех этапов: подготовительного, бескислородного (гликолиз, анаэробное дыхание) и кислородного (аэробное дыхание). У многих многоклеточных животных связан с пищеварительной, дыхательной и кровеносной системами.
Подготовительный этап энергетического обмена
Происходит в цитоплазме клеток всех организмов, в желудочно-кишечном тракте у большинства многоклеточных животных и человека. Под действием ферментов большие органические молекулы расщепляются на мономеры. Эти процессы происходят с выделением незначительного количества энергии, которое рассеивается в виде тепла.
Бескислородный (анаэробный) этап энергетического обмена
Происходит в клетках, всегда предшествует аэробному у большинства организмов (способных использовать кислород).
Анаэробное расщепление – это простейшая известная форма образования и аккумулирования энергии в макроэргических связях молекул АТФ. Суть его состоит в расщеплении молекулы глюкозы преимущественно путем гликолиза на две молекулы пировиноградной или молочной кислоты (особенно в мышечных клетках). Две молекулы пировиноградной кислоты (С3Н403) при определенных условиях могут восстанавливаться до молочной (С3Н603). Суммарное уравнение гликолиза:
С6Н1206 + 2АДФ + 2Н3Р04→2С3Н603 + 2АТФ + 2Н20.
Во время гликолиза выделяется около 200 кДж энергии, часть которой расходуется на синтез двух молекул АТФ (84 кдж), а часть рассеивается в виде тепла (116 кДж).
Процесс гликолиза энергетически малоэффективный, так как в макроэргических связях АТФ аккумулируется лишь 35-40 % энергии. Это связано с тем, что не происходит полного распада веществ. Конечные продукты гликолиза еще содержат в себе много энергии в химических связях.
Гликолиз имеет чрезвычайно большое физиологическое значение, несмотря на его низкую эффективность. В условиях дефицита кислорода организм благодаря гликолизу может получать энергию. И вдобавок конечные продукты – пировиноградная и молочная кислоты – в аэробных условиях подвергаются дальнейшему ферментативному расщеплению.
Некоторые микроорганизмы и беспозвоночные животные (преимущественно паразиты) являются анаэробами и не могут использовать кислород. Им присущ лишь анаэробный энергетический обмен.
Существует несколько типов преобразования глюкозы, органических соединений без доступа кислорода с аккумуляцией энергии в виде АТФ, которые называются брожением. Известны спиртовое брожение (у некоторых дрожжей и бактерий с образованием спирта), маслянокислое (с образованием масляной кислоты), молочнокислое (у молочнокислых бактерий с образованием молочной кислоты) и т. п.
Суммарное уравнение спиртового брожения:
С6Н1206 + 2АДФ + 2Н3Р04→2С2Н5ОН + 2АТФ + 2Н20 + 2С02.
Кислородный этап энергетического обмена (аэробное дыхание)
Происходит в митохондриях. Органические соединения, которые образовались в бескислородном этапе, окисляются до конечных продуктов (углекислого газа и воды). Соединения окисляются с отщеплением от них водорода. С помощью веществ-переносчиков он передается кислороду с образованием воды. Этот процесс называется тканевым дыханием. При этом выделяется большое количество энергии, которое аккумулируется в связях АТФ. В кислородном этапе можно выделить реакции цикла Кребса и те, что протекают на дыхательной цепи.
Значение и функции
Этот восьмиэтапный циклический процесс, итогом которого является окисление ацетильного остатка до углекислого газа, может показаться излишне сложным. Тем не менее, он имеет огромное значение в метаболизме промежуточных реакций и выполняет ряд функций. К ним относятся:
- энергетическая;
- анаболическая;
- катаболическая;
- транспортная.
Цикл Кребса участвуют в катаболизме жиров и углеводов. Соединения, образующиеся на разных стадиях процесса, участвуют в синтезе многих необходимых для организма веществ — глутамина, порфиринов, глицина, фенилаланина, цистеина и других. Когда промежуточные продукты покидают ЦК для участия в синтезе, происходит их замещение с помощью так называемых анаплеротических реакций, которые катализируются регуляторными ферментами, например, пируваткарбоксилазой.
Транспортная функция ЦК заключается в содействии гликолизу. Глюкозу невозможно превратить сразу в АТФ, поэтому механизм гликолиза действует поэтапно и сопровождается постоянным перемещением атомов и катионов водорода от одних соединений к другим. Для их транспортировки нужны специальные соединения, которые получаются на одной из стадий ЦТК. Участвующие в гликолизе коферменты цикла Кребса:
- НАД*H+(Никотинамидадениндинуклеотид с катионом водорода). Образуется на III стадии ЦК.
- ФАД*H2 (Флавинадениндинуклеотид с молекулой водорода). Появляется на V стадии ЦК.
Реакции ЦК имеют и большое клиническое значение. Хотя для людей не свойственны мутации, связанные с генами ферментов, участвующих в цикле, однако их редкие проявления губительны для здоровья. Они могут приводить к опухолям мышц и почек, нарушениям работы нервной системы.
Предыдущая
БиологияСтроение микроскопа — схема устройства с обозначениями и подписями частей
Следующая
БиологияСистематика в биологии — основы классификации растений и животных
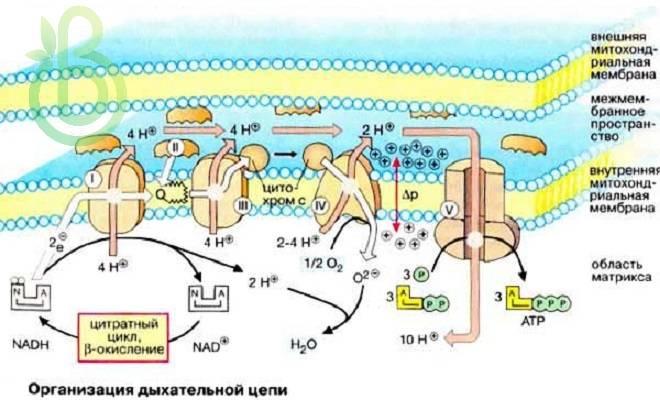
Строение ферментативных комплексов дыхательной цепи
1 комплекс. НАДН-КоQ-оксидоредуктаза
Этот комплекс также имеет рабочее название НАДН-дегидрогеназа, содержит ФМН, 42 белковых молекулы, из них не менее 6 железосерных белков.
Функция
- Принимает электроны от НАДН и передает их на коэнзим Q (убихинон).
- Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
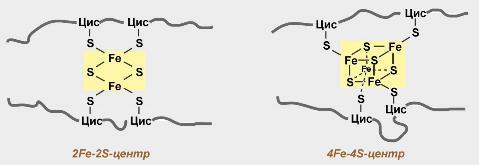
Железосерные белки (FeS-белки) – это белки содержащие атомы железа, которые соединены с атомами серы и с серой остатков цистеина. В результате образуется железо-серный центр.
2 комплекс. ФАД-зависимые дегидрогеназы
Данный комплекс как таковой не существует, его выделение условно. К нему относятся ФАД-зависимые ферменты, расположенные на внутренней мембране – например, ацил-SКоА-дегидрогеназа (β-окисление жирных кислот), сукцинатдегидрогеназа (цикл трикарбоновых кислот), митохондриальная глицерол-3-фосфат-дегидрогеназа (челночный механизм переноса атомов водорода).
Функция
- Восстановление ФАД в окислительно-восстановительных реакциях.
- Обеспечение передачи электронов от ФАДН2 на железосерные белки внутренней мембраны митохондрий. Далее эти электроны попадают на коэнзим Q (убихинон).
3 комплекс. КоQ-цитохром c-оксидоредуктаза
По другому данный комплекс называется цитохром средуктаза. В его составе имеются молекулы цитохрома b и цитохрома c1, железо-серные белки. Комплекс представляет собой 2 мономера, в каждом из которых насчитывается 11 полипептидных цепей.
Функция
- Принимает электроны от коэнзима Q и передает их на цитохром с.
- Переносит 2 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
Имеются разногласия по поводу количества переносимых ионов H+ при участии 3-го и 4-го комплексов. По одним данным, третий комплекс переносит 2 иона H+ и четвертый комплекс переносит 4 иона H+. По другим авторам, наоборот, третий комплекс переносит 4 иона H+ и четвертый комплекс переносит 2 иона H+.
4 комплекс. Цитохром с-кислород-оксидоредуктаза
В этом комплексе находятся цитохромы а и а3, он называется также цитохромоксидаза, состоит из 13 субъединиц. В комплексе имеются ионы меди, соединенные с белками комплекса через HS-группы цистеина, и формирующие центры, подобные тем, что имеются в железо-серных белках.
Функция
- Принимает электроны от цитохрома с и передает их на кислород с образованием воды.
- Переносит 4 иона Н+ на наружную поверхность внутренней митохондриальной мембраны.
5 комплекс
Пятый комплекс – это фермент АТФ-синтаза, состоящий из множества белковых цепей, подразделенных на две большие группы:
- одна группа формирует субъединицу Fo (произносится со звуком «о», а не «ноль» т.к олигомицин-чувствительная) – ее функция каналообразующая, по ней выкачанные наружу протоны водорода устремляются в матрикс.
- другая группа образует субъединицу F1 – ее функция каталитическая, именно она, используя энергию протонов, синтезирует АТФ.
Механизм работы АТФ-синтазы получил название вращательный катализ.
Упрощенно считают, что для синтеза 1 молекулы АТФ необходимо прохождение через АТФ-синтазу приблизительно 3-х ионов Н+, и еще один ион H+ используется для транспорта неорганического фосфата в матрикс митохондрии и для антипорта АДФ (в митохондрию) АТФ (в цитозоль). Таким образом, для получения одной молекулы АТФ из АДФ требуется 4 иона Н+.
Поскольку при прохождении одной пары электронов по всей дыхательной цепи выкачивается 10 ионов Н+, то окисление одного моля НАДН+Н+ приведет к образованию 2,5 молей АТФ.
Метаболизм
Одно из важнейших свойств живого (вспоминаем биологию) — это обмен веществ с окружающей средой. Действительно, только живое существо может что-то поглощать из окружающей среды, и что-то потом в неё выделять.
В биохимии обмен веществ принято называть «метаболизм». Обмен веществ, обмен энергией с окружающей средой — это метаболизм.
Когда мы, допустим, съели бутерброд с курицей, мы получили белки (курица) и углеводы (хлеб). В процессе пищеварения белки распадутся до аминокислот, а углеводы — до моносахаров. То, что я описал сейчас, называется катаболизм, то есть распад сложных веществ на более простые. Первая часть метаболизма — это катаболизм.
Ещё один пример. Ткани в нашем организме постоянно обновляются. Когда отмирает старая ткань, её обломки растаскивают макрофаги, и они заменяется новой тканью. Новая ткань создаётся в процессе синтеза белка из аминокислот. Синтез белка происходит в рибосомах. Создание нового белка (сложного вещества) из аминокислот (простого вещества) — это анаболизм.
Итак, анаболизм — это противоположность катаболизму. Катаболизм — это разрушение веществ, анаболизм — это создание веществ. Кстати, чтобы их не путать, запомните ассоциацию: «Анаболики. Кровью и потом». Это голливудский фильм (довольно скучный, на мой взгляд) о спортсменах, применяющих анаболики для роста мышц. Анаболики — рост, синтез. Катаболизм — обратный процесс.
Метаболизм веществ
С пищей в организм поступают три основные группы сложных биохимических соединений — белки, жиры и углеводы. Они являются первичными метаболитами, потому что участвуют в обмене веществ или в метаболизме. Этот процесс происходит между любыми живыми клетками и окружающей средой непрерывно. Суть цикла Кребса заключается в том, что он является областью схождения двух путей метаболизма. Это следующие процессы:
- катаболизм, при котором происходит распад более сложных веществ на простые, в частности, глюкозы на моносахариды;
- анаболизм — синтез сложных веществ из простых, например, белков из аминокислот.
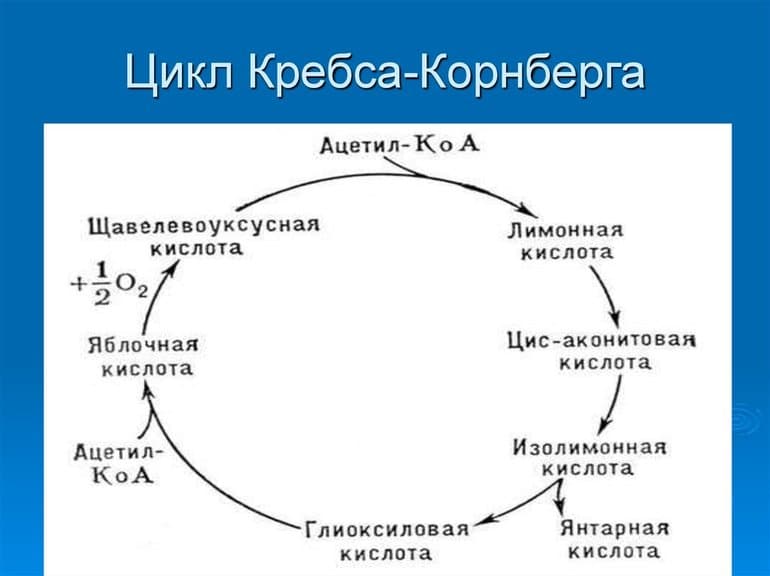
После попадания в пищеварительную систему сложные вещества расщепляются под действием ферментов на более простые, которые внутри клеток превращаются сначала в пируват (пировиноградную кислоту), а затем — в ацетильный остаток. Все эти преобразования можно назвать подготовкой к ЦК, а образование остатка — его запуском или начальным этапом.
Дальнейшие стадии цикла трикарбоновых кислот являются частью катаболизма. Процесс идет каскадно. Каждый предыдущий этап запускает последующий, а промежуточные продукты химических реакций служат не только для продолжения цикла, но и при определенных потребностях организма могут пополнять запасы веществ, необходимых для синтеза новых соединений (анаболизма).
https://youtube.com/watch?v=4Y_PAnqz2LM
Регуляция цикла Кребса[править | править код]
В разных тканях цикл Кребса выполняет разные функции. Так, в мышцах и головном мозге в цикле Кребса ацетил-КоА окисляется с образованием НАДН и ФАДН2, которые используются для синтеза АТФ в дыхательной цепи. В печени при голодании ацетил-КоА не окисляется в цикле Кребса. Вместо этого цикл Кребса используется для синтеза малата из некоторых аминокислот, который превращается в оксалоацетат и вступает в глюконеогенез. После приема пищи ацетил-КоА в печени и жировой ткани поступает в цикл Кребса на очень короткий период. В первой реакции цикла он преобразуется в цитрат, затем экспортируется в цитозоль и используется для биосинтеза жирных кислот.
Изоцитратдегидрогеназаправить | править код
Изоцитратдегидрогеназа ингибируется при высокой концентрации НАДН. Кофермент изоцитратдегидрогеназы — НАД+. Когда НАД+ восстанавливается до НАДН, изоцитратдегидрогеназа инактивируется, поскольку ее кофермент отсутствует.
Клеточное дыхание
Для нормальной жизнедеятельности живым клеткам постоянно требуется энергия. Ее главный универсальный источник — аденозинтрифосфат (АТФ), способный встраиваться в белки организма напрямую. Это соединение получается в результате ряда реакций окисления, носящих общее название «клеточное дыхание». При этом происходит постепенный распад органических веществ вплоть до простейших неорганических — углекислого газа CO2 и воды H2O.
Структурное строение молекул АТФ содержит фосфорангидридные связи, которые имеют свойство накапливать высвобожденную при прохождении реакций клеточного дыхания энергию, поэтому называются макроэргическими. Так создаются энергетические запасы клеток, которые могут высвобождается при необходимости разрывом этих связей. Процесс синтеза АТФ и класса вспомогательных соединений включает три этапа:
Гликолиз происходит в цитоплазме.
В матриксе митохондрий проходят все химические реакции цикла Кребса.
Окислительное фосфорилирование на внутренней мембране митохондрий.
Преобразование аденозиндифосфата (АДФ) в АТФ характерно для всех этапов. Но наибольшее суммарное количество молекул с макроэргическими связями образуется при фосфорилировании. Это не значит, что процессы гликолиза и ЦК менее важны. Многие соединения, образующиеся во время их протекания, участвуют в регуляции клеточного дыхания.
Химизм
На конечном этапе цикла ключевой фермент цикла АТФ-цитратлиаза разрушает цитрат на оксалоацетат и ацетил-CoA, используя энергию одной АТФ. Далее происходят следующие реакции:
- Ацетил~CoA + CO2 + ФдH2 → Пируват + Фд
- Пируват + АТФ → ФЕП + АДФ + Фн
- ФЕП + АТФ + 2H+ → глицеральдегид-3-фосфат + АМФ + ФФн
Полученный глицеральдегид-3-фосфат используется для синтеза материалов клетки. Общее уравнение ассимиляции имеет вид:
- 3CO2 + 2ФдH2 + 2НАДФН·H+ + ФАДH2 + НАДН·Н+ + 5АТФ→ глицеральдегид-3-фосфат
Ассимиляция углекислого газа идёт с большой затратой АТФ и восстановительных эквивалентов. В целом цикл считается анаэробным, поскольку включает много ферментов и восстановителей (Fe4S4 бактериальных ферредоксинов), которые чувствительны к кислороду (из аэробных бактерий такой цикл обнаружен только у Hydrogenobacter).
Этот цикл является возможным кандидатом в реакции, которые могли протекать на Земле в добиологических условиях, и, по этой причине, изучение этого цикла представляет интерес для понимания происхождения жизни. Принципиально, что данный цикл является автокаталитическим — в нём, в отличие от окислительного ЦТК в результате одного оборота цикла количество реагентов, в том числе акцепторов СО2, увеличивается, а не уменьшается, как в цикле Кребса. Так же, было доказано, что некоторые из реакций можно катализировать, используя обычные минералы, в частности пять реакций протекает в присутствии ZnS (сфалерит) на свету.
Ферменты
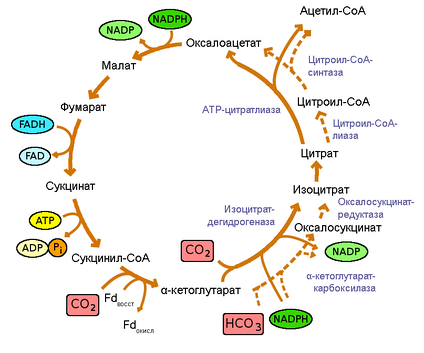
Цикл Арнона. Пунктиром обозначены альтернативные пути реакции.
Считается, что три реакции цикла Кребса необратимы, а значит, в цикле Арнона эти реакции должны осуществляться другими ферментами, чтобы сделать цикл обратимым: так, сукцинатдегидрогеназа заменена на фумаратредуктазу, НАД+-зависимая α-кетоглутаратдегидрогеназа на ферредоксин-зависимую α-кетоглутаратсинтазу, а цитратсинтаза на АТФ-цитратлиазу. Эти три фермента обычно считаются характерными для данного цикла.
Большинство бактерий, которые используют этот цикл — мезофилы, однако представители филума Aquificae растут при температуре 70 °C и выше (Aquifex aeolicus вплоть до 95 °C). При таких условия термически неустойчивый распадается на сукцинат и коэнзим А, что сопровождается потерей энергии и делает невозможным его дальнейшее использование в цикле. Aquificae (как было показано на ) решают эту проблему, тратя дополнительный АТФ на превращение α-кетоглутарата в изоцитрат совместным действием биотин-зависимой α-кетоглутараткарбоксилазы и недекарбоксилирующей изоцитратдегидрогеназы, что делает процесс необратимым при повышенной температуре.
У некоторых видов основной путь цикла может иметь некоторые отличия. Так вместо АТФ-цитратлиазы расщепление цитрата могут осуществлять два других фермента: цитрил-CoA синтаза и . Однако, оба эти фермента филогенетически близки к АТФ-цитратлиазе. Их обнаружили у представителей Aquificaceae (но не у других Aquificae). Более того, некоторые протеобактерии (например, Magnetococcus sp. штамм MC-1), вероятно, обладают новым типом АТФ-цитратлиазы.