Сухожильный орган гольджи
Содержание:
- Функции
- Функции комплекса Гольджи
- АНАТОМИЯ НЕРВНО-МЫШЕЧНОГО ВЕРЕТЕНА
- Феноменальная сила. Обратная сторона
- Строение аппарата Гольджи
- Мышечные веретена
- Комплекс (аппарат) Гольджи
- Строение мышечного веретена
- Виды рецепторов
- Строение аппарата Гольджи
- Функции комплекса Гольджи
- Потенциал действия мышечного волокна
- Строение
- Образование лизосом
Функции
Аппарат Гольджи часто считается отделом распределения и доставки химических веществ клетки. Он модифицирует белки и липиды (жиры), которые продуцируются в эндоплазматическом ретикулуме, и готовит их для экспорта за пределы клетки или для транспортировки в другие места внутри клетки. Белки и липиды, построенные в гладком и шероховатом эндоплазматическом ретикулуме, укладываются в крошечные пузырьковые везикулы, которые движутся через цитоплазму, пока не достигнут комплекса Гольджи.
Везикулы сливаются с мембранами Гольджи и высвобождают, содержащиеся внутри молекулы в органеллу. Оказавшись внутри, соединения дополнительно обрабатываются с помощью аппарата Гольджи, а затем направляются в везикуле к месту назначения внутри или вне клетки. Экспортируемые продукты представляют собой секреции белков или гликопротеинов, которые являются частью функции клетки в организме. Другие вещества возвращаются в эндоплазматический ретикулум или могут созревать, чтобы впоследствии стать лизосомами.
Модификации молекул, которые осуществляются в комплексе Гольджи, происходят упорядоченным образом. Каждая цистерна имеет два основных отдела: цис-отдел — это конец органеллы, где вещества поступают из эндоплазматического ретикулума для обработки, а также транс-отдел, где они выходят в форме меньших отдельных везикул. Следовательно, цис-отдел расположен вблизи эндоплазматического ретикулума, откуда поступает большая часть веществ, а транс-отдел расположен вблизи плазматической мембраны клетки, куда отправляются многие из веществ, модифицирующиеся в аппарате Гольджи.
Химический состав каждого отдела, а также ферменты, содержащиеся в люменах (внутренних открытых пространствах цистерн) между отделами, являются отличительными. Белки, углеводы, фосфолипиды и другие молекулы, образующиеся в эндоплазматическом ретикулуме, переносятся на аппарат Гольджи, чтобы подвергнутся биохимическому модифицированию при переходе от цис к транс-отделам комплекса. Ферменты, присутствующие в люмене Гольджи, модифицируют углеводную часть гликопротеинов путем добавления или вычитания отдельных мономеров сахара. Кроме того, аппарат Гольджи сам по себе производит самые разнообразные макромолекулы, включая полисахариды.
Комплекс Гольджи в растительных клетках продуцирует пектины и другие полисахариды, необходимые для структуры растений и обмена веществ. Продукты, экспортируемые аппаратом Гольджи через транс-отдел, в конечном итоге сливаются с плазматической мембраной клетки. Среди наиболее важных функций комплекса — сортировка большого количества макромолекул, продуцируемых клеткой, и их транспортировка в необходимые пункты назначения. Специализированные молекулярные идентификационные метки или метки, такие как фосфатные группы, добавляются ферментами Гольджи, чтобы помочь в этом процессе сортировки.
Функции комплекса Гольджи
Протеолиз белковых веществ, что приводит к активации белков, так проинсулин переходит в инсулин.
Обеспечивает транспорт из клетки продуктов синтеза ЭПС.
Синтез полисахаридов, таких как пектин, гемицеллюлоза, которые входят в состав мембран растительных клеток, образование гликозаминогликанов, одного из составляющих межклеточной жидкости.
В цистернах пластинчатого комплекса идет созревание белковых веществ, необходимых для секреции, трансмембранных протеинов клеточной мембраны, ферментов лизосом и др. В процессе созревания белки постепенно перемещаются по отделам органоида, в которых завершается их формирование и происходит гликозилирование и фосфорилирование.
Формирование липоптротеидных веществ. Синтез и накопление слизистых веществ (муцина). Образование гликолипидов, которые входят в состав мембранного гликокаликса.
Передает белки в трех направлениях: к лизосомам (перенос контролируется ферментом – маннозой- 6-фосфат), к мембранам или внутриклеточной среде, и к межклеточному пространству.
Вместе с зернистой ЭПС образует лизосомы, путем слияния отпочковавшихся везикул с автолитическими ферментами.
Экзоцитозный перенос – везикула, подойдя к мембране, встраивается в нее и оставляет свое содержимое с наружной стороны клетки.
АНАТОМИЯ НЕРВНО-МЫШЕЧНОГО ВЕРЕТЕНА
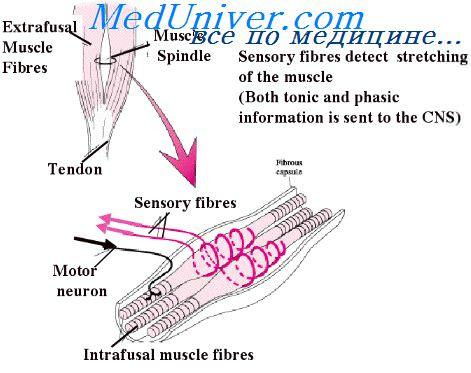
Нервно-мышечное веретено – это совокупность проприоцепторов, расположенных в скелетных мышцах, отслеживающих изменения в длине тканей. Мышечные волокна, находящиеся внутри богато иннервированной сенсорными нервными окончаниями соединительнотканной капсулы и входящие в состав веретен, называются интрафузальными.
Сенсорные, или афферентные мышечные волокна, отслеживают напряжение и степень растяжения мышцы. Если растяжение слишком интенсивно или происходит слишком быстро, альфа-афферентные волокна воспринимают это как потенциальную угрозу целостности мышцы, вынуждая экстрафузальные (то есть лежащие за пределами соединительнотканной капсулы) волокна сокращаться, укорачивая мышцу, защищая её таким образом от чрезмерного растягивания. Этот ответ называется миотатическим рефлексом.
Если вы когда-нибудь бывали на приёме у невролога, вы наверняка наблюдали этот рефлекс в действии. Врач ударяет молоточком по надколенному сухожилию, что приводит к сокращению мышцы-разгибателя бедра, что вызывает непроизвольное разгибание голени – это сигнал, что нервно-мышечное веретено в норме.
Все скелетные мышцы имеют нервно-мышечные веретена, защищающие их от повреждения миотатическим рефлексом. Помимо немедленной реакции, заключающейся в сокращении мышцы в ответ на возможность повреждения тканей, нервно-мышечное веретено способно проводить перекалибровку самого себя, подстраиваясь под стимул. Если экстрафузальные волокна изменяют свою длину, защищая мышцу, то гамма-эфферентные волокна снижают или повышают активность нервно-мышечного веретена, обеспечивая мониторинг длины мышцы. Это позволяет нервно-мышечному веретену быстрее реагировать на изменение длины мышцы. У вас когда-нибудь дрожали мышцы, например, при выполнении планки или работе со штангой? Именно так проявляется в этом случае активность мышечных веретен.
Феноменальная сила. Обратная сторона
Возможно и самопроизвольное отключение тормозящего сухожильного рефлекса в стрессовой ситуации. Вы могли слышать о подобных историях, когда, например, не занимающийся спортом человек вдруг приподнимает автомобиль, чтобы спасти зажатого ребенка, или перескакивает с разбега через двухметровый забор, спасаясь от собак.
Не рекомендую Вам пытаться вызвать подобный эффект целенаправленно. Регулярные тренировки с субмаксимальными или статическими нагрузками в зале не столь героичны, зато намного более безопасны для Вашего организма. Ведь в случае опасности организм рискует здоровьем ради спасения своей жизни или кого-то из близких, и только в таких случаях это оправдано.
Строение аппарата Гольджи
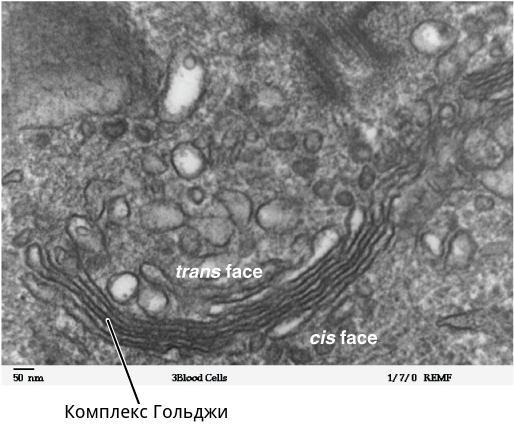
Комплекс Гольджи по внешнему виду, видимому в электронный микроскоп, напоминает стопку наложенных друг на друга дискообразных мешочков, около которых находится множество пузырьков. Внутри каждого «мешка» находится узкий канал, расширяющийся на концах в так называемые цистерны (иногда цистерной называют весь мешочек). От них отпочковываются пузырьки. Вокруг центральной стопки формируется система взаимосвязанных трубочек.
С наружней, имеющей несколько выпуклую форму, стороны стопки образуются новые цистерны путем слияния пузырьков отпочковывающихся от гладкой эндоплазматической сети. На внутренней стороне цистерны завершают свое созревание и распадаются снова на пузырьки. Таким образом, цистерны (мешочки стопки) Гольджи перемещаются от наружней стороны к внутренней.
Часть комплекса, располагающаяся ближе к ядру, называется «цис». Та, что ближе к мембране, – «транс».
Микрофотография комплекса Гольджи
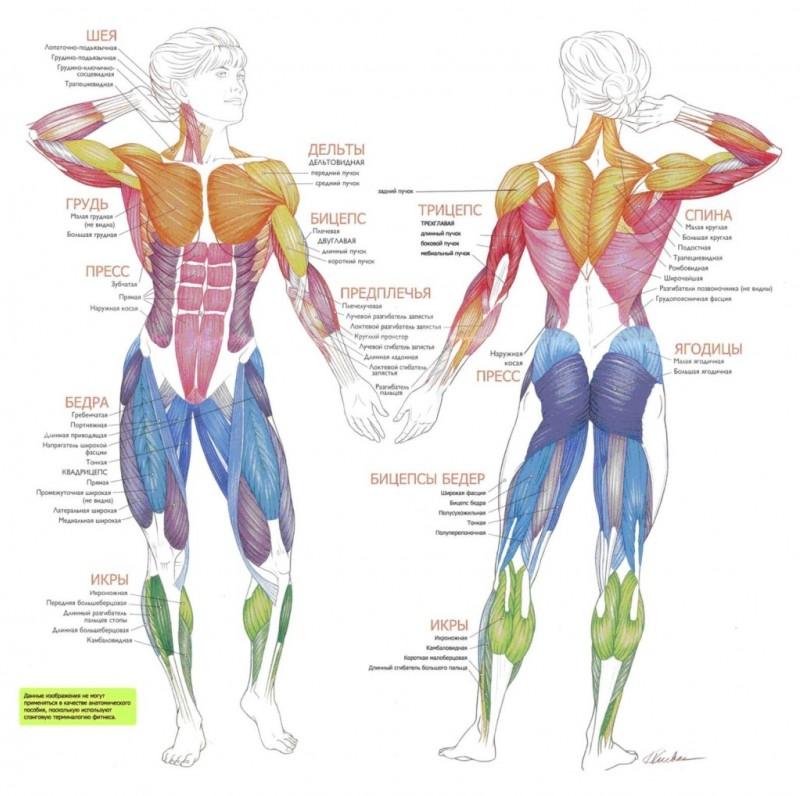
Мышечные веретена
Мышечные веретена – механорецепторы, неравномерно распределяемые в теле животных и человека.
В отдельных мышцах, таких как экстраокулярные мышцы у кошки и кролика они вообще отсутствуют (встречаются в аналогичных мышцах у человека). Всего 1-2 веретена обнаружены в мелких мышцах хвоста крысы.
В крупных мышцах человека их может быть до нескольких сот. Наибольшая плотность обнаружена в небольших мышцах рук, совершающих тонкие высококоординированные движения.
Расположение мышечных веретен таково, что они включены как бы параллельно основным экстрафузальным волокнам скелетной мышцы. При сокращении экстрафузальных мышц интрафузальные волокна с окружающими их нервными волокнами испытывают меньшую нагрузку, т.е. разгружаются.
Сухожильные органы, напротив, расположены последовательно с экстрафузальными волокнами и при их сокращении испытывают возрастающую нагрузку.
Интрафузальные волокна получают моторную иннервацию, благодаря которой они активно участвуют в деятельности мышечных рецепторов. Собственно рецептирующими элементами веретена являются окончания тонких нервных волоконец, оплетающих структуры интрафузальных мышц в их центральной зоне.
Основу веретена составляют пучки параллельно распложенных интрафузальных волокон. Число волокон, их размер может сильно варьировать. Так у рептилий имеется всего одно волокно, у земноводных (лягушки) – 3-12, у млекопитающих (кошки) – 2-13.
Диаметр волокон может колебаться от 6 до 28 мкм, при этом в составе даже одного пучка могут оказаться как толстые, так и тонкие волокна. Длина волокон может достигать нескольких миллиметров.
Волокна, входящие в состав веретена, могут начинаться от разных экстрафузальных волокон, но сходятся они у дистального конца. В 50-х годах 20 века было впервые обнаружено существование двух типов веретен. Обнаруженные различия между веретенами относились к строению экваториальной области интрафузальных волокон.
В полярных областях интрафузальных волокон хорошо видны ядра, рассеянные в основном по периферии волокна, как это имеет место в обычных экстрафузальных мышечных волокнах.
Это и послужило основанием называть центральную зону волокна областью ядерной сумки. Следует заметить, что никакой специальной сумки для ядер, иной, чем сарколемма самого волокна, нет. Волокна такого типа получили наименование волокон с ядерной сумкой – ЯС-волокон.
У волокон другого типа, которые в большинстве случаев являются более тонкими и короткими, концентрация ядер в экваториальной области меньше, и они располагаются в ряд, образуя цепочки. Такие волокна получили название волокон с ядерной цепочкой – ЯЦ-волокон.
У некоторых животных, например, кроликов, ЯЦ-волокна отсутствуют. У ЯС-волокон экваториальная зона отделена от остального волокна переходной областью (область миотрубки). Какой-либо резкой границы между миотрубкой и обычными частями интрафузального волокна нет.
У волокон обоих типов центральная ядерная область невелика, достигает всего лишь 300 мкм (у человека длина 100-250 мкм), а при растяжении – 500 мкм. В обоих случаях миофибриллы в центральной области истончаются и образуют очень тонкий слой. В ЯЦ-волокнах миофибриллы содержатся в большем количестве, чем в ЯС-волокнах.
Вблизи экваториальной области отмечается наличие эластической ткани. Число ЯЦ- и ЯС-волокон сильно варьирует в разных мышцах и у разных животных. Например, в крупных веретенах человека, где общее число волокон достигает 14, 3-4 волокна относятся к ЯС-волокнам, а остальные – к ЯЦ-волокнам.
ЯЦ-волокна чаще встречаются у животных, обладающих более тонкой координацией движений.
Центральная область обычных волокон, входящих в состав одного веретена, окружена капсулой. Величина ее может достигать у человека 5 мм. ЯС-волокна и длинные ЯЦ-волокна могут простираться далеко за пределы капсулы. Наибольшего размера (80-200 мкм) капсула достигает в средней части. Упругость капсулы, так же как и интрафузальных волокон, увеличивается за счет эластических волокон.
Однако во внутрикапсулярной жидкости обнаружено большое количество кислых мукополисахаридов. Такое отличие в составе лимфы и жидкости капсулы заставляет предположить, что гиалуроновые кислоты образуются оболочкой веретена.
Таким образом, можно полагать, что между лимфатическим пространством и внутрикапсулярным пространством имеется диффузный барьер, затрудняющий проникновение веществ через капсулу веретен.
Комплекс (аппарат) Гольджи
Как и в любой другой клетке, в мышечном волокне имеется органелла общего назначения, известная под названием комплекса или аппарата Гольджи. Эту органеллу обнаружил в 1889 году итальянский ученый Камилло Гольджи. В саркоплазме мышечного волокна таких органелл насчитывается тысячи.
Строение комплекса Гольджи
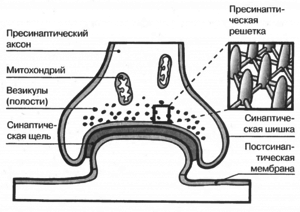
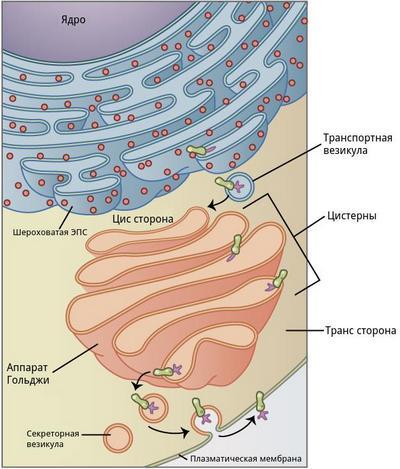
Комплекс Гольджи представляет собой стопку дискообразных мембранных цистерн (рис.1) и связанную с ними систему пузырьков (везикул). Между цистернами имеются связывающие их трубчатые структуры. 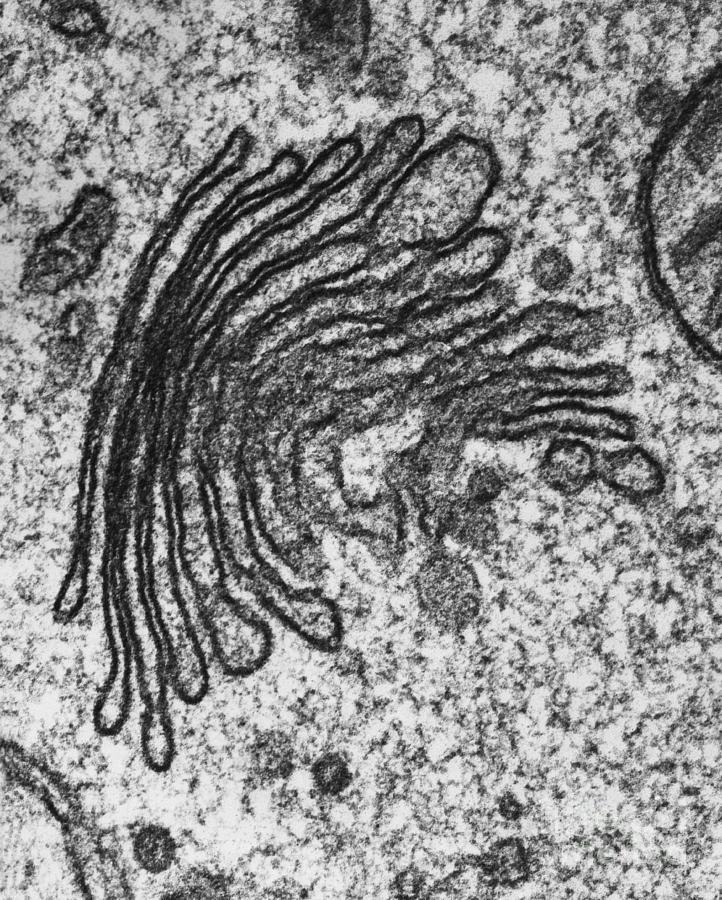
Рис.1. Комплекс Гольджи
Комплекс Гольджи расположен недалеко от ядер мышечного волокна и шероховатой эндоплазматической сети (рис.2).
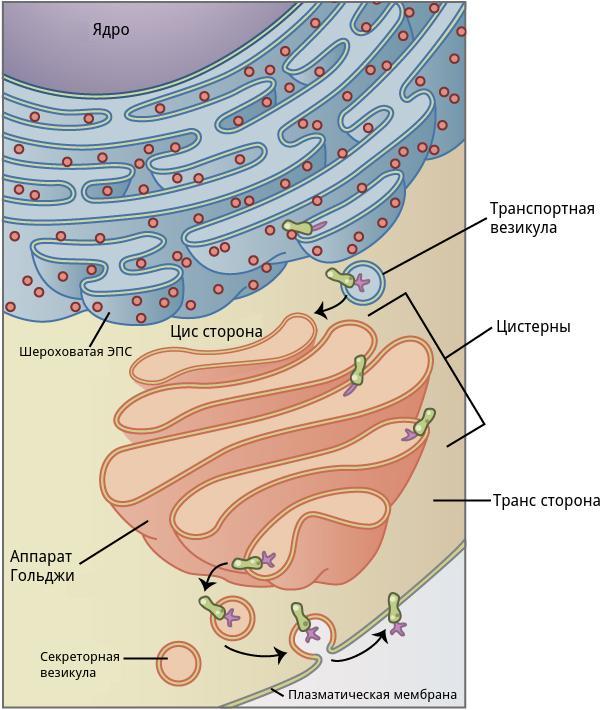
Рис.2. Структура и функции комплекса Гольджи
В комплексе Гольджи различают три отдела: цис-сеть Гольджи, некомпактные зоны и транс-сеть Гольджи.
Цис-сеть Гольджи расположена перед первой цистерной, ближе к ядру мышечного волокна. В транс-сеть Гольджи, переходит последняя цистерна комплекса. Она расположена на более удаленном расстоянии от ядра мышечного волокна. Некомпактные зоны расположены между соседними стопками аппарата Гольджи.
Перенос и преобразование белков
Необходимые мышечному волокну белки синтезируются на рибосомах. Затем они перемещаются в шероховатую эндоплазматическую сеть. Эти белки называют «незрелыми», потому что они еще не приняли свою пространственную структуру (то есть не свернулись определенным образом). Из шероховатой эндоплазматической сети белки перемещаются в виде мембранных пузырьков в цис-сеть комплекса Гольджи. Известно, что в цистернах, располагающихся ближе к ядру мышечного волокна, содержатся наименее зрелые белки. В цистернах аппарата Гольджи белки «созревают», то есть приобретают свою трехмерную структуру. Биохимики называют этот этап процессингом белка. Каким образом созревающие белки перемещаются по цистернам комплекса Гольджи до сих пор непонятно.
Более подробно строение и функции мышц описаны в моих книгах «Гипертрофия скелетных мышц человека» и «Биомеханика мышц«
В конце концов от цистерн, расположенных достаточно далеко от ядер отпочковываются пузырьки, содержащие полностью «зрелые» белки. С помощью мембранных пузырьков эти белки доставляются «по адресу» в зависимости от полученных ими в аппарате Гольджи «меток». Образно говоря, в мышечном волокне комплекс Гольджи играет роль почты, в которой сортируются и оправляются по адресам различные «посылки» в виде белков, заключенных в пузырьки.
Сборка мембран
В комплексе Гольджи происходит также сборка мембран. Вещества, из которых состоят мембраны (белки, липиды) поступают в комплекс Гольджи из эндоплазматической сети. Затем в цистернах комплекса Гольджи собираются участки мембран, из которых изготавливаются мембранные пузырьки. Они перемещаются в саркоплазме мышечного волокна в те места, где нужно достроить мембрану.
Транспорт веществ к клеточной мембране
Часть веществ, синтезированных в мышечном волокне, выводится наружу. Эти вещества накапливаются в комплексе Гольджи, упаковываются в мембранные пузырьки и транспортируются к сарколемме и выводятся за её пределы.
Образование лизосом
В комплексе Гольджи также образуются лизосомы – органеллы мышечного волокна, играющие существенную роль в катаболизме белков.
Строение мышечного веретена
Структура и функции мышечных веретен очень сложны. Они присутствуют в
большинстве
скелетных мышц
, особенно их много в мышцах, требующих тонкой регуляции движений
(например, в мелких мышцах кисти). Что касается крупных мышц, то мышечных
веретен больше всего в мышцах, содержащих много
медленных фазических волокон (волокон I типа; slow twitch fibers)
.
Форму мышечного (или нервно-мышечного) веретена характеризует само
название этой структуры; веретено состоит из пучка модифицированных
мышечных волокон, иннервируемых и сенсорными, и двигательными аксонами (
рис. 38.1
). Его диаметр равен примерно 100 мкм, длина — до 10 мм. Иннервированная
часть мышечного веретена заключена в соединительнотканную капсулу. Так
называемое лимфатическое пространство капсулы заполнено жидкостью.
Мышечное веретено свободно располагается между обычными мышечными
волокнами. Его дистальный конец прикреплен к
эндомизию
— соединительнотканной сети внутри мышцы. Мышечные веретена лежат
параллельно обычным
поперечно-полосатым мышечным волокнам
Такое расположение важно с функциональной точки зрения.
Мышечное веретено содержит модифицированные мышечные волокна, называемые
интрафузальными мышечными волокнами
в отличие от обычных —
экстрафузальных мышечных волокон
. Интрафузальные волокна гораздо тоньше, чем экстрафузальные, и слишком
слабы, чтобы участвовать в сокращении мышцы. Различают два типа
интрафузальных мышечных волокон: с ядерной сумкой и с ядерной цепочкой (
рис. 38.2
). Их названия связаны с организацией клеточных ядер. Волокна с ядерной
сумкой крупнее, чем с ядерной цепочкой, и их ядра плотно упакованы в
средней части волокна наподобие сумки с апельсинами. В волоконах с ядерной
цепочкой все ядра расположены в один ряд.
Мышечные веретена получают сложную иннервацию. Сенсорная иннервация
состоит из одного
афферентного аксона группы Iа
и нескольких
афферентов группы II
(
рис. 38.2
). Афференты группы Iа относятся к классу сенсорных аксонов наибольшего
диаметра со скоростью проведения от 72 до 120 м/с; аксоны группы II имеют
меньший, чем Iа, диаметр и проводят импульсы со скоростью от 36 до 72 м/с
(
табл. 33.2
). Афферентный аксон группы Iа образует первичное окончание, спирально
обвивающее каждое интрафузальное волокно
Первичные окончания есть на
интрафузальных волокнах обоих типов, что важно для деятельности этих
рецепторов. Афференты группы II образуют вторичные окончания на волокнах с
ядерной цепочкой.
Двигательную иннервацию мышечных веретен обеспечивают два типа
гамма-эфферентных аксонов
(
рис. 38.2
). Динамические гамма-эфференты оканчиваются на каждом волокне с ядерной
сумкой, статические гамма- эфференты — на волокнах с ядерной цепочкой.
Гамма-эфферентные аксоны тоньше, чем альфа-эфференты экстрафузальных
мышечных волокон, поэтому они проводят возбуждение с меньшей скоростью (
табл. 33.3
).
Виды рецепторов
Существуют несколько классификаций рецепторов:
- По положению в организме
- Экстерорецепторы (экстероцепторы) — расположены на поверхности или вблизи поверхности тела и воспринимают внешние стимулы (сигналы из окружающей среды)
- Интерорецепторы (интероцепторы) — расположены во внутренних органах и воспринимают внутренние стимулы (например, информацию о состоянии внутренней среды организма)
- По способности воспринимать разные стимулы
- Мономодальные — реагирующие только на один тип раздражителей (например, фоторецепторы — на свет)
- Полимодальные — реагирующие на несколько типов раздражителей (например, многие болевые рецепторы, а также некоторые рецепторы беспозвоночных, реагирующие одновременно на механические и химические стимулы)
- По адекватному раздражителю:
- Хеморецепторы — воспринимают воздействие растворённых или летучих химических веществ
- Осморецепторы — воспринимают изменения осмотической концентрации жидкости (как правило, внутренней среды)
- Механорецепторы — воспринимают механические стимулы (прикосновение, давление, растяжение, колебания воды или воздуха и т. п.)
- Фоторецепторы — воспринимают видимый и ультрафиолетовый свет
- Терморецепторы — воспринимают понижение (холодовые) или повышение (тепловые) температуры (тепловые стимулы)
- Болевые рецепторы, стимуляция которых приводит к возникновению болевых ощущений. Такого физического стимула, как боль, не существует, поэтому выделение их в отдельную группу по природе раздражителя в некоторой степени условно. В действительности, они представляют собой высокопороговые сенсоры различных (химических, термических или механических) повреждающих факторов. Однако уникальная особенность ноцицепторов, которая не позволяет отнести их, например, к «высокопороговым терморецепторам», состоит в том, что многие из них полимодальны: одно и то же нервное окончание способно возбуждаться в ответ на несколько различных повреждающих стимулов.
- Электрорецепторы — воспринимают изменения электрического поля
- Магнитные рецепторы — воспринимают изменения магнитного поля
У человека имеются первые шесть типов рецепторов. На хеморецепции основаны вкус и обоняние, на механорецепции — осязание, слух и равновесие, а также ощущения положения тела в пространстве, на фоторецепции — зрение. Терморецепторы есть в коже и некоторых внутренних органах. Большая часть интерорецепторов запускает непроизвольные и в большинстве случаев неосознаваемые, вегетативные рефлексы. Так, осморецепторы включены в регуляцию деятельности почек, хеморецепторы, воспринимающие pH, концентрации углекислого газа и кислорода в крови, включены в регуляцию дыхания и т. д.
Иногда предлагается выделять группу электромагнитных рецепторов, в которую включают фото-, электро- и магниторецепторы. Магниторецепторы точно не идентифицированы ни у одной группы животных, хотя предположительно ими служат некоторые клетки сетчатки птиц, а возможно, и ряд других клеток.
В таблице приведены данные о некоторых типах рецепторов
| Природа раздражителя | Тип рецептора | Место расположения и комментарии |
|---|---|---|
| • электрическое поле | • ампула Лоренцини и другие типы | • Имеются у рыб, круглоротых, амфибий, а также у утконоса и ехидны |
| • химическое соединение | • хеморецептор | |
| • влажность | • гигрорецептор | • Относятся к осморецепторам или механорецепторам. Располагаются на антеннах и ротовых органах многих насекомых |
| • механическое воздействие | • механорецептор | • У человека имеются в коже (экстероцепторы) и внутренних органах (барорецепторы, проприоцепторы) |
| • давление | • барорецептор | • Относятся к механорецепторам |
| • положение тела | • проприоцептор | • Относятся к механорецепторам. У человека это нервно-мышечные веретена, сухожильные органы Гольджи и др. |
| • осмотическое давление | • осморецептор | • В основном интерорецепторы; у человека имеются в гипоталамусе, а также, вероятно, в почках, стенках желудочно-кишечного тракта, возможно, в печени. Существуют данные о широком распространении осморецепторов во всех тканях организма |
| • свет | • фоторецептор | |
| • температура | • терморецептор | • Реагируют на изменение температуры. У человека имеются в коже и в гипоталамусе |
| • повреждение тканей | • ноцицептор | • В большинстве тканей с разной частотой. Болевые рецепторы — свободные нервные окончания немиелинизированных волокон типа C или слабо миелинизированных волокон типа Aδ. |
| • магнитное поле | • магнитные рецепторы | • Точное расположение и строение неизвестны, наличие у многих групп животных доказано поведенческими экспериментами |
Строение аппарата Гольджи
Если мы посмотрим на аппарат Гольджи в электронный микроскоп, то увидим него нечто напоминающее стопку наложенных друг на друга мешочков, около которых находится множество пузырьков. В середине каждого подобного мешка находится узкий канал, который расширяется на концах в так званые цистерны. От них в свою очередь отпочковываются пузырьки. Вокруг центральной стопки образуется система связанных между собой трубочек.
Внешняя сторона аппарат Гольджи имеет немного выпуклую форму, там наши стопки образуют новые цистерны путем слияния пузырьков отпочковывающихся от гладкой эндоплазматической сети. С внутренней стороны аппарата цистерны завершают свое созревание и также распадаются вновь на пузырьки. Подобным образом происходит перемещение цистерн (мешочков, стопок) от наружной стороны органеллы к внутренней.
Также часть комплекса Гольджи, которая располагается ближе к ядру клетки, называется «цис», а часть, которая находится ближе к мембране, называется «транс».
Так выглядит аппарат Гольджи на рисунке.
Функции комплекса Гольджи
Роль аппарата Гольджи в жизни клетки разнообразна, в основном она сводится к модификации и перераспределению синтезирующих веществ и также их выведению за пределы клетки, образованию лизосом и построению цитоплазматической мембраны.
Весьма высока активность аппарата Гольджи в секреторных клетках. Белки, которые поступающие из эндоплазматической сети концентрируются в аппарате Гольджи, затем в пузырьках Гольджи переносятся к мембране.
В клетках растений при формировании клеточной стенки именно Гольджи секретирует углеводы, которые служат матриксом для нее. При помощи микротрубочек отпочковавшиеся пузырьки Гольджи перемещаются и их мембраны сливаются с цитоплазматической мембраной, а содержимое включается в клеточную стенку.
Комплекс Гольджи бокаловидных клеток (они находятся в толще эпителия слизистой оболочки кишечника и дыхательных путей) секретирует гликопротеин муцин, он образует слизь.
А в клетках кишечника именно аппарат Гольджи выполняет важную функцию по перемещению липидов. Происходит это таким образом: жирные кислоты и глицерол попадают в клетки, затем в эндоплазматической сети происходит синтез своих липидов, большая часть их которых покрывается белками и при помощи Гольджи транспортируется к клеточной мембране, пройдя через которую липиды окажутся в лимфе.
Также благодаря аппарату Гольджи происходит формирование лизосом, на которых более детально остановимся в будущей статье.
Потенциал действия мышечного волокна
Природа и механизм возникновения потенциала действия рассмотрены в главе 5. ПД МВ длится 1–5 мс, скорость его проведения по сарколемме, включая T-трубочки, составляет 3–5 м/с.
Орлов Р.С., Ноздрачёв А.Д. Нормальная физиология : Учебник. — М.: ГЭОТАР-Медиа, 2009. – 688 с. Глава 7. Физиология сократительных элементов. — Скелетная мышца. — Иннервация. — Двигательная иннервация. — С. 101-102.
Экстрафузальное мышечное волокно (истор.; лат. extra вне, снаружи + гист. fusus веретено) — общее название поперечнополосатых мышечных волокон, составляющих основную массу скелетных мышц и обладающих способностью к сокращению на всем протяжении. (Словарь медицинских терминов)/
2] Интрафузальное мышечное волокно (син. волокно мышечное внутриверетенное — myofibra intrafusalis, LNH) мышечное волокно, входящее в состав нервно-мышечного веретена; периферические части интрафузального мышечного волокна обладают поперечной исчерченностью и способны к сокращению, центральная часть не сокращается.
Шванн Теодор (Schwann T.), немецкий гистолог и физиолог (1810–1882); вместе с М. Шульце создал клеточную теорию (1839 г.); в 1836 г. открыл пепсин, в 1838 г. опубликовал первую работу по строению миелиновой оболочки.
Строение
Аппарат Гольджи образован системой пузырьков, трубочек и мембранных взаимосвязанных мешочков – цистерн. Мембранная цистерна – структурная и функциональная единица, называемая диктиосомой. Органоид находится рядом с ядром клетки или клеточным центром.
Рис. 1. Комплекст Гольджи.
У растений, простейших и некоторых беспозвоночных животных органелла представляет собой рассеянные мелкие образования и пузырьки. Комплекс Гольджи у животных (позвоночных) сосредоточен в одном месте и имеет форму плотных мембранных стопок, соединённых трубочками.
Аппарат Гольджи состоит из трёх отделов, в каждом из которых располагается свой набор ферментов. Подробно об отделах и ферментах органеллы представлено в таблице.
|
Отдел |
Расположение |
Ферменты |
Функции |
|
Цис-отдел |
Самый близкий к ядру |
Фосфогликозидаза |
Присоединяет фосфат к моносахариду |
|
Медиальный |
Промежуточный между двумя крайними отделами. Состоит из нескольких мембранных стопок |
Манназидаза |
Отщепляет моносахарид |
|
N-ацетилглюкозаминтрансфераза |
Присоединяет специфичные углеводы |
||
|
Транс-отдел |
Максимально отдалён от ядра |
Пептидаза |
Участвует в гидролизе белков |
|
Трансфераза |
Переносит группы химических соединений |
Через аппарат Гольджи проходят мембранные белковые пузырьки (везикулы), отделяющиеся от промежуточной эндоплазматической сети (ЭПС). Незрелые белки попадают в цис-отдел, где разделяются на две группы. В медиальном отделе белки проходят модификацию. В транс-отделе происходит синтез и разделение (сегрегация) веществ по функциональному признаку.
Образование лизосом
Многие гидролитические ферменты лизосом проходят через аппарат Гольджи, где они получают «метку» в виде специфического сахара — маннозо-6-фосфата (М6Ф) — в составе присоединённого к аминокислотной цепочке олигосахарида. Добавление этой метки происходит при участии двух ферментов. Фермент N-ацетилглюкозаминфосфотрансфераза специфически опознает лизосомальные гидролазы по деталям их третичной структуры и присоединяет N-ацетилглюкозаминфосфат к шестому атому нескольких маннозных остатков олигосахарида гидролазы. Второй фермент — фосфогликозидаза — отщепляет N-ацетилглюкозамин, создавая М6Ф-метку. Затем эта метка опознается белком-рецептором М6Ф, с его помощью гидролазы упаковываются в везикулы и доставляются в лизосомы. Там, в кислой среде, фосфат отщепляется от зрелой гидролазы. При нарушении работы N-ацетилглюкозаминфосфотрансферазы из-за мутаций или при генетических дефектах рецептора М6Ф все ферменты лизосом «по умолчанию» доставляются к наружной мембране и секретируются во внеклеточную среду. Выяснилось, что в норме некоторое количество рецепторов М6Ф также попадают на наружную мембрану. Они возвращают случайно попавшие во внешнюю среду ферменты лизосом внутрь клетки в процессе эндоцитоза.