Миотатический рефлекс: значение и физиология
Содержание:
- Сенсомоторные кожные рефлексы
- Ib афференты: Golgi Tendon Organ reflex
- Как понять, что рефлекс присутствует?
- Двигательные рефлексы[править | править код]
- Развитие рефлекторной деятельности
- Исторические сведения
- Нарушение мышечного тонуса и отсутствие рефлекса
- Обратный миотатический рефлекс
- Реакция первичного окончания мышечного веретена. Рефлекс на растяжение мышцы
- Литература
- Патологические рефлексы
- Активная модуляция рефлексов
- Group II афференты. Снова веретена
Сенсомоторные кожные рефлексы
Сенсомоторные, или кожные, рефлексы заключаются в том, что в ответ на стимуляцию кожи происходит сокращение мышц. Если, например, провести заостренным предметом по коже живота, то брюшные мышцы сократятся (абдоминальный рефлекс). В отличие от проприоцептивных рефлексов, в данном случае рецептор и эффектор разнесены и расположены в разных органах. Более того, в таких рефлексах в рефлекторную дугу входит несколько синапсов и несколько вставочных нейронов, поэтому их часто называют полисинаптическими рефлексами. Вставочные нейроны позволяют вовлекать в рефлекторную реакцию соседние сегменты спинного мозга и контралатеральную сторону тела.
Полисинаптические рефлексы характеризуются более продолжительным латентным периодом (например, 70-150 мс для мигательного рефлекса глазного века), быстрой утомляемостью и аккомодацией, а также таким явлением, как суммация подпороговых стимулов. Данный термин описывает инициацию рефлекса постоянным повторением слабых стимулов, каждый из которых в отдельности не вызвал бы рефлекторной реакции. Например, длительное раздражение слизистой оболочки носа суммируется, постепенно достигает порога, и мы, наконец, чихаем. В числе других примеров подобных защитных рефлексов можно упомянуть кашель, выделение слез и такие пищевые рефлексы, как глотание и сосание.
Ib афференты: Golgi Tendon Organ reflex
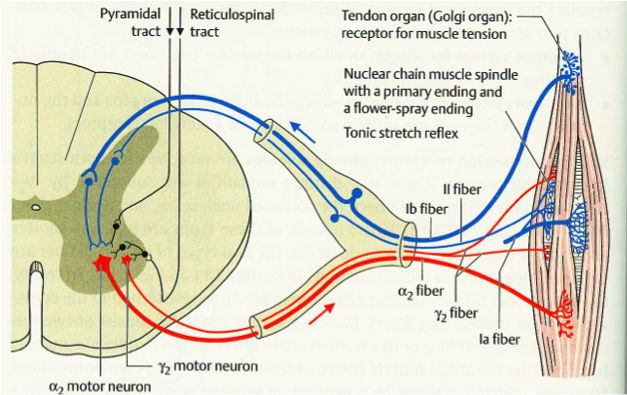
Смена концепций
Когда-то считалось, что GTO реагирует лишь на сильное натяжение мышцы, и выполняет защитный рефлекс аутогенного торможения. Это утверждение полностью опровергнуто, начиная с 1967 года, когда было показано, что даже активация единственной моторной единицы (motor unit) активирует GTO, если соответствующие волокна к нему присоединены.
Тем не менее, по всей видимости именно интернейроны Ib афферентов выполняют функции защиты диапазона движения, но совместно с суставными рецепторами (Circ.Spinal.Cord.p454).
Структура
Рефлекс от проприоцепторов GTO дисинаптический. Сигнал от Ib афферентов достигает интернейронов (которые называют Ib интернейроны, по их доминирующему входу), а не моторных нейронов, и они уже затем что-то делают. В силу дисинаптической природы этой рефлексной цепи, это «что-то» может быть совсем разным, поэтому описывать его стоит словом «может»
- Этот рефлекс может тормозить альфа-мотонейроны этой же мышцы, а также синергичные мышцы
- Этот рефлекс может возбуждать альфа-мотонейроны мышц-антагонистов. В этих двух функциях GTO рефлекс противоположен по действию миотатическому рефлексу. Степень «может» определяется привходящими влияниями, супра- и интра-спингальными.
- Этот рефлекс может наоборот способствовать возбуждению мышцы, если она несет нагрузку, и должна ее нести: например, «автогенное торможение» сменяется возбуждением в фазе разгибания/опоры в локомоции.
Некоторые особенности
Несмотря на то, что Ib вход для них является основным, Ib интернейроны получают влияния и от Ia и II входов тоже (в меньшей степени).
Схема GTO рефлекса не подвержена влиянию возвратного торможения.
Однако тактильные ощущения от кожи (cutaneous volleys) способны подавить тормозящий эффект GTO рефлекса, эффект называется cutaneous suppression, и продемонстрировано на разгибателях ноги в ответ на стимуляцию подошвы (Circ.Spinal.Cord.p227).
В других комбинациях кожные сигналы способны, наоборот, усилить GTO рефлекс.
Вообще, Ib интернейроны имеют тенденцию менять знак при увеличении силы входа: например, вестибулярные сигналы (от гальваностимуляции) усиливают GTO рефлекс, когда они слабые, и тормозят, когда они становятся сильнее. Такой же эффект описан для кортикоспинальных влияний. (Circ.Spinal.Cord.p232).
Как понять, что рефлекс присутствует?

Очень важно знать, присутствует ли у мышцы данный рефлекс. Чтобы это определить, как правило, используют кинезиологический тест
Для того чтобы его провести, важно присутствие двух человек, иначе будет сложно сделать тест.
С самого начала нужно понять, что за мышцу тестируют, а также какие функции она выполняет в обычной жизни. Необходимо проследить за движением именно этой мышцы. Миотатический рефлекс будет заметен, если не обращать внимания на работу других мышц. Далее мышцы проверяют с помощью дополнительной нагрузки и смотрят за тем, как она себя поведет. В этом пункте главное не сила, а способность мышц перестраиваться. Чтобы это посмотреть, нужно изометрическим способом напрячь мышцу, то есть должно быть препятствие, в которое можно будет упереться со всем давлением. По мере повышения данной нагрузки можно легко увидеть, есть миотатический рефлекс или нет. Если мышцы выдерживают и реагируют нормально, то он присутствует, в противном случае – его нет. Как правило, роль препятствия выполняет человек, который проводит тест. Именно по этой причине одного человека мало.
Двигательные рефлексы[править | править код]
Для понимания спортивно-прикладных аспектов двигательной деятельности человека в рамках теории рефлексов наиболее важны рефлексы, определяющие особенности мышечной работы спортсмена. Это, прежде всего, безусловные двигательные рефлексы спинного мозга, к которым относятся рефлексы на растяжение мышцы, кожные сгибательные рефлексы удаления от раздражителя, рефлексы отталкивания, ритмические, позные рефлексы. Спинной мозг обусловливает также взаимодействие центров мышц-антагонистов (в том числе реципрокную иннервацию, рационально регулирующую одновременную работу мышц-агонистов и антагонистов), а при локомоциях — более сложные разгибательные и шагательные рефлексы (описание условных рефлексов в данном случае опускается).
Миотатический рефлекс — рефлекс на натяжение мышц, проявляющийся в форме возбуждения мышцы в ответ на ее натяжение. Он играет кардинально важную роль при выполнении множества произвольных, в том числе спортивных, в особенности мощностных, движений, поскольку только натянутая мышца развивает, при прочих равных условиях, наибольшее усилие. Эффектами, обусловленными миотатическим рефлексом, обусловливаются такие важнейшие закономерности работы мышц, как взаимосвязь «длина-напряжение» (чем сильнее, до оптимума, натянута мышца, тем большее усилие она развивает), «скорость-сила» (чем быстрее натяжение мышцы, тем больше развиваемая сила; чем быстрее сокращение мышцы, тем развиваемая ею сила меньше), а также различные режимы работы мышечного аппарата.
Шейно-тонический рефлекс — физиологический феномен, обусловливающий взаимосвязь между движениями (установкой) головы относительно туловища и тонусом мышц туловища и рук. В традиционной физиологии (школа Р. Магнуса) он рассматривается как явление, в норме характерное только для животных и младенцев, но во взрослом состоянии (за исключением патологии) легко подавляемое и поэтому якобы несущественное. Однако многолетняя практика спорта и специальные исследования показывают весьма значительную роль шейно-тонического рефлекса в управлении двигательными действиями спортсмена и их влияние на мышечную силу и координацию движений. Конкретное действие шейно-тонического рефлекса проявляется в виде зависимости, при которой движение (или фиксированная установка) головы с ее сгибанием (наклоном вперед), разгибанием (наклоном назад) или наклоном в сторону стимулируют деятельность одноименных мышц туловища и особенно мышц плечевого пояса. При исполнении и, в особенности, освоении спортивных упражнений эффекты шейно-тонического рефлекса могут оказывать существенное, а часто решающее влияние на координацию двигательных действий и потому широко используются в качестве одного из средств формирования двигательного навыка и технического совершенствования произвольного движения.
Развитие рефлекторной деятельности
Развитие рефлекторной деятельности в эмбриогенезе проходит несколько стадий: 1. Стадия первичных двигательных Р. (первые Р. у эмбрионов человека обнаруживаются на 3-м месяце пренатального развития и проявляются в виде сгибания шеи, движения частей туловища и конечностей при раздражении губ и крыльев носа, а осуществление реакций обеспечивается рефлекторными дугами, проходящими через ядра тройничного нерва в продолговатом мозгу и через шейное утолщение спинного мозга). 2. Стадия первичной генерализации Р. (реакции проявляются в виде быстрых движений головы, туловища и конечг о-стей). 3. Стадия вторичной генерализации Р. (рефлексогенной зоной становится практически любой участок тела, а в двигательные реакции вовлекаются большие группы мышц; т. о. наблюдается широкая иррадиация возбуждения в ц, н. с., к-рая в более позднем периоде эмбрионального развития понижается и Р. становятся более ограниченными и специализированными). 4. Стадия специализации Р. (формируются сложные пищевые — сосательный, глотательный, защитные — чесатель-ный, отряхивательный и установочные — шейные, лабиринтные Р.). Очень рано у человеческого плода обнаруживаются подошвенный, коленный и хватательный Р.
В постнатальном периоде происходит быстрая консолидация готовых при рождении Р., а также формирование новых в пределах развивающихся анализаторных систем. Дальнейшая координация рефлекторных актов происходит за счет созревания синаптических структур и механизмов центрального торможения (см.). В более поздние сроки постнатального периода формируются такие сложные специализированные реакции, как, напр., ориентировочно-исследовательская реакция (см.).
Различные возрастные периоды человека характеризуются также различиями в степени сформированностп в. н. д. и стабильностью основных врожденных Р. По мере старения всего организма в нервных клетках происходят необратимые структурные изменения, снижающие их функциональные возможности (падение возбудимости, уменьшение скорости проведения возбуждения). Развивается инертность процессов в ц. н. с., ослабляются тормозные процессы, увеличивается время рефлекторных реакций.
Исследование Р. проводят с помощью специальных рефлексометров, позволяющих определять основные параметры и степень проявления рефлекторных реакций (см. Рефлексометрия). В клин, практике основные соматические и вегетативные Р. человека оценивают визуально при нанесении раздражений на различ-иые рефлексогенные зоны. Техника вызывания Р. состоит в нанесении удара перкуторным молоточком по сухожилию или надкостнице; раздражение кожи, слизистых оболочек осуществляют специальными предметами (тупая игла, волосковая кисточка, бумажная полоска). Для анализа центральных механизмов осуществления Р. используют широко применяемые электрофизиологические методы исследования.
Исторические сведения
Понятие о рефлексе впервые возникло в физике Декарта. Декарт развивал общую механическую картину мира и хотел включить в неё также и поведение живых существ. Концепция Декарта была создана в эпоху, когда разные учёные давали материалистические объяснения феноменам, происходящим в природе: например, медик Уильям Гарвей открыл и описал систему кровообращения, в которой эта система выступала как простой механизм с известными к тому времени человечеству элементами — насосом, «трубами» и т. п. Теория Декарта ещё более укрепляла принцип материалистического детерминизма, так как Гарвей рассматривал в качестве механизма только внутреннее устройство тела животного, а Декарт перенёс этот принцип также и на взаимодействие организмов с внешним миром, то есть, по сути, на психическую деятельность. Декарт считал, что взаимодействие организмов с окружающими телами опосредовано нервной машиной, в которой мозг выполняет роль центра, а от него расходятся «нервные трубки». Согласно его схеме, внешние факторы действуют на концы расположенных в теле нервных «нитей», которые, натягиваясь, открывают клапаны отверстий, ведущих из мозга в нервы. По эвного акта нервной системы».
Предположение о полностью рефлекторном характере деятельности высших отделов головного мозга впервые было развито ученым-физиологом И. М. Сеченовым. До него физиологи и неврологи не решались поставить вопрос о возможности физиологического анализа психических процессов, которые предоставлялось решать психологии.
Далее идеи И. М. Сеченова получили развитие в трудах И. П. Павлова, который открыл пути объективного экспериментального исследования функций коры, разработал метод выработки условных рефлексов и создал учение о высшей нервной деятельности. Павлов в своих трудах ввёл деление рефлексов на безусловные, которые осуществляются врождёнными, наследственно закреплёнными нервными путями, и условные, которые, согласно взглядам Павлова, осуществляются посредством нервных связей, формирующихся в процессе индивидуальной жизни человека или животного.
Большой вклад в формирование учения о рефлексах внёс Чарлз С. Шеррингтон (Нобелевская премия по физиологии и медицине, 1932). Он открыл координацию, взаимное ингибирование и облегчение рефлексов.
Нарушение мышечного тонуса и отсутствие рефлекса
Тонус мышц – это обычный процесс, который предполагает самопроизвольную активацию всех мышц, причем реакция протекает без участия мозговой деятельности, она происходит сама по себе. Это является нормой функционирования всех рефлексов тела человека.
Основная роль тонических скелетных мышц заключается в обеспечении поддержки мышц, которые благодаря этому всегда готовы к активации, то есть к напряжению. Также тонус влияет на такой важный момент, как поддержание любого выбранного положения тела. Тонус не создает тяжести, даже когда человек находится в сидячем или ином положении.
Важно то, что происходят различные нарушение мышечного тонуса, а, следовательно, и миотатического рефлекса, неодинаково. Этот процесс строго индивидуален
Необходимо, чтобы тонус был средним, так как слишком пониженный влияет на работоспособность и активность, человек не выдерживает даже слабые физические и умственные нагрузки. При сильном, чрезмерном тонусе почти невозможно сосредоточиться на работе, человек находится в постоянном напряжении и не может повлиять на этот процесс.
Обратный миотатический рефлекс
Активация
сухожильных органов Гольджи
сопровождается рефлекторной реакцией, которая, на первый взгляд,
противоположна
рефлексу на растяжение
(на самом деле, как мы увидим ниже, эта реакция дополняет его). Реакция
называется обратным миотатическим рефлексом; соответствующая рефлекторная
дуга представлена на
рис. 38.12
. Сенсорные рецепторы этого рефлекса — сухожильные органы Гольджи в прямой
мышце бедра.
Афферентные аксоны
входят в
спинной мозг
, разветвляются и образуют синаптические окончания на интернейронах. Путь
от сухожильных органов Гольджи не имеет моносинаптической связи с
альфа-мотонейронами
, а включает в себя тормозные интернейроны, подавляющие активность
альфа-мотонейронов прямой мышцы бедра, и возбуждающие интернейроны, которые
вызывают активность альфа- мотонейронов мышц-антагонистов.
Таким образом, по своей организации обратный миотатический рефлекс
противоположен рефлексу на растяжение, откуда и произошло его название.
Однако в действительности обратный миотатический рефлекс функционально
дополнителен рефлексу на растяжение. Сухожильные органы Гольджи служат
датчиком силы, развиваемой
сухожилием
, с которым они соединены. Когда при поддержании стабильной позы
(например, человек стоит в положении «смирно») прямая мышцы бедра
начинает утомляться, прилагаемая к коленному сухожилию сила уменьшается и,
следовательно, снижается активность соответствующих сухожильных органов
Гольджи. Поскольку обычно эти рецепторы подавляют активность
альфа-мотонейронов прямой мышцы бедра, ослабление импульсных разрядов от
них приводит к повышению их возбудимости и сила, развиваемая мышцей,
возрастает. В итоге происходит координированное изменение рефлекторных
реакций с участием как
мышечных веретен
, так и афферентных аксонов сухожильных органов Гольджи; сокращение прямой
мышцы усиливается, и поза сохраняется.
При чрезмерной активации рефлексов можно наблюдать
рефлекс «складного ножа»
. Когда сустав пассивно сгибается, сопротивление сначала увеличивается.
Однако по мере дальнейшего сгибания сопротивление внезапно падает и сустав
резко переходит в свое конечное положение. Причина этого — рефлекторное
торможение. Раньше рефлекс «складного ножа» объясняли активацией
сухожильных рецепторов Гольджи, поскольку считалось, что у них высокий
порог реакции на мышечное растяжение. Однако теперь рефлекс связывают с
активацией других высокопороговых мышечных рецепторов, находящихся в
мышечной фасции.
Реакция первичного окончания мышечного веретена. Рефлекс на растяжение мышцы
Когда длина мышечного веретена изменяется внезапно, первичное окончание (но не вторичное) стимулируется особенно мощно. Это избыточное возбуждение первичного окончания называют динамическим ответному что означает чрезвычайно активную реакцию первичного окончания на высокую скорость изменения длины веретена. Даже когда длина веретена увеличивается лишь на долю микрометра и это увеличение осуществляется в течение доли секунды, первичный рецептор передает громадное количество дополнительных импульсов по большим сенсорным нервным волокнам диаметром 17 мкм, но только до тех пор, пока длина действительно увеличивается. Как только увеличение длины прекращается, этот дополнительный прирост импульсного разряда возвращается к гораздо более низкому уровню, чем статический разряд, все еще присутствующий в ответе.
Наоборот, при укорочении веретена происходит прямо противоположное изменение сигнала. Таким образом, первичное окончание посылает чрезвычайно сильные, положительные или отрицательные, сигналы к спинному мозгу, информируя его о любом изменении длины мышечного веретена.
Регуляция интенсивности статических и динамических ответов гамма-мотонейронами. Гамма-двигательные нервы к мышечному веретену можно разделить на два типа: гамма-динамические (гамма-d) и гамма-статические (гамма-s). Первые из них возбуждают главным образом интрафузальные волокна с ядерной сумкой, а вторые возбуждают в основном интрафузальные волокна с ядерной цепочкой. Когда гамма-d волокна возбуждают волокна с ядерной сумкой, динамический ответ мышечного веретена становится чрезвычайно усиленным, тогда как статический ответ почти не меняется.
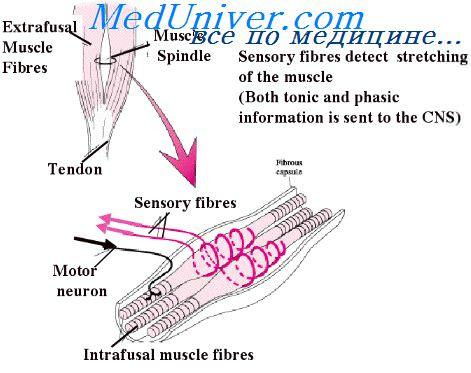
Наоборот, стимуляция гамма-s волокон, возбуждающих мышечные волокна с ядерной цепочкой, усиливает статический ответ, оказывая лишь незначительное влияние на динамический ответ.
Непрерывный разряд мышечных веретен при нормальных условиях. В норме, особенно на фоне некоторой степени возбуждения гамма-эфферентных волокон, в сенсорных нервных волокнах мышечных веретен постоянно возникают импульсы. Растяжение мышечных веретен увеличивает частоту импульсов, тогда как укорочение веретен снижает ее. Таким образом, веретена могут посылать к спинному мозгу положительные сигналы, т.е. увеличенное количество импульсов, свидетельствующее о растяжении мышцы, или отрицательные сигналы, т.е. количество импульсов ниже нормы, что указывает на отсутствие растяжения мышцы.
Рефлекс на растяжение мышцы
Самым простым проявлением функции мышечных веретен является рефлекс на растяжение мышцы. Всякий раз, когда мышца внезапно растягивается, возбуждение веретен вызывает рефлекторное сокращение крупных мышечных волокон растянутой мышцы и тесно связанных с ней мышц-синергистов.
Нервный контур рефлекса на растяжение. На рисунке изображен основной контур рефлекса на растяжение мышечных веретен. Видно, что проприоцептивное нервное волокно типа 1а, исходящее от мышечного веретена, входит в задний корешок спинного мозга. Затем ветвь этого волокна идет непосредственно к переднему рогу серого вещества спинного мозга и синаптически связывается с передними мотонейронами, которые посылают двигательные нервные волокна к той же мышце, от которой исходят волокна мышечных веретен. Таким образом, существует моносинаптический путь, позволяющий рефлекторному сигналу вернуться с кратчайшей задержкой назад к мышце после возбуждения веретена. Большинство волокон типа II от мышечных веретен заканчиваются на множестве вставочных нейронов серого вещества, и их аксоны проводят сигналы к передним мотонейронам с задержкой или выполняют другие функции.
— Также рекомендуем «Динамические и статические рефлексы на растяжение. Демпфирующий механизм сокращения»
Оглавление темы «Физиология спинальной рефлекторной деятельности»:1. Реакция первичного окончания мышечного веретена. Рефлекс на растяжение мышцы2. Динамические и статические рефлексы на растяжение. Демпфирующий механизм сокращения3. Гамма-эфферентная система мышечного сокращения. Стабилизация положения тела4. Рефлекс на растяжение в клинике. Клонус мышечных волокон5. Сухожильный рефлекс Гольджи. Значение сухожильного рефлекса6. Функция мышечных веретен. Сгибательный рефлекс и механизм отдергивания7. Перекрестный разгибательный рефлекс. Реципрокное торможение и иннервация8. Позные и локомоторные рефлексы спинного мозга. Шагательные движения9. Шагание на месте. Чесательный рефлекс и рефлексы мышечного спазма10. Спинальные вегетативные рефлексы. Спинальный шок
Литература
- Скоромец А. А., Скоромец А. П., Скоромец Т. А. Пропедевтика клинической неврологии. СПб.: Политехника, 2004
- Косицкий Г. И., «Физиология человека». Изд. «Медицина», 1985.
- Словарь физиологических терминов / отв. ред. Газенко О.Г.. — М.: «Наука», 1987. — 32 000 экз.
- Фундаментальная и клиническая физиология: Учебник для студентов высших учебных заведений / ред. Камкин А.Г., Каменский А.А.. — М.: Издательский центар «Академия», 2004. — 1072 с. — 5 000 экз. — ISBN 5-7695-1675-5.
- Ноздрачев А.Д., Баранникова И.А., Батуев А.С. и др. Физиология нервной, мышечной и сенсорной систем // Общий курс физиологии человека и животных. В 2 кн.: Учеб. для биол. и мед. спец. вузов. — М.: «Высшая школа», 1991. — Т. 1. — 512 с. — 48 000 экз. — ISBN 5-06-000650-6.
- Сергеев Б.Ф. Ступени эволюции интеллекта. — М.: Наука, 1986. — 192 с.
Патологические рефлексы
Основная статья: Пирамидные знаки
Патологические рефлексы — неврологический термин, обозначающий необычные для здорового взрослого человека рефлекторные реакции. В ряде случаев свойственны более ранним стадиям фило- или онтогенеза.
Рефлексы орального автоматизма являются одним из проявлений псевдобульбарного синдрома, который возникает при двустороннем поражении пирамидных путей, идущих к двигательным ядрам языкоглоточного, блуждающего, добавочного и подъязычного нервов (каудальной группы черепных нервов). К ним относятся: Хоботковый рефлекс, Дистанс-оральный рефлекс Карчикяна, Назо-лабиальный рефлекс Аствацатурова, Ладонно-подбородочный рефлекс Маринеску-Радовичи (Marinescu-Radovici), «Бульдожий» рефлекс Янышевского.
К патологическим кистевым рефлексам относятся: рефлекс Тремнера, Якобсона-Ласка, Жуковского, Бехтерева запястно-пальцевой, верхний рефлекс Россолимо, большепальцевой сгибательный рефлекс Клиппеля—Вейля.
Патологические стопные рефлексы делятся на разгибательные и сгибательные. К первым относятся: рефлекс Оппенгейма, рефлекс Гордона, рефлекс Шеффера, рефлекс Бабинского, рефлекс Чаддока, рефлекс Бинга, рефлекс Штрюмпеля. Ко вторым относятся: нижний рефлекс Россолимо, Рефлекс Бехтерева-Менделя, Рефлекс Жуковского-Корнилова, Рефлекс Бехтерева, Рефлекс Пуусеппа.
Одним из патогномоничных признаков поражения пирамидного пути являются клонусы. Проявляются быстрыми и ритмичными сокращениями мышцы или группы мышц в ответ на их растяжение. Клонус стопы — вызывается у больного, лежащего на спине, а клонус надколенной чашечки — у больного, лежащего на спине с выпрямленными ногами.
Рефлекторное содружественное движение конечности, сопутствующее произвольному движению другой конечности называется Синкинезией. Синкинезии могут быть как физиологическими (например, размахивание руками во время бега или быстрой ходьбы), так и патологическими. Патологические синкинезии подразделяют на 3 типа: глобальные синкинезии, имитационная синкинезия, координаторная синкинезия. Синкинезии развиваются не в остром, а в отдалённом периоде основного заболевания, приведшего к поражению пирамидных путей. Их патогенез не до конца изучен. Они часто воспринимаются больными со спинальной травмой как признак восстановления нервной проводимости спинного мозга.
Активная модуляция рефлексов
Модуляция моноаминами
Спинальные рефлексы подвержены модуляции моноаминами (применительно к спинному мозгу и моторным цепям имеют в виду серотонин, норадреналин и дофамин). Такая модуляция экспериментально продеменострирована как сильная для группы II и вообще FRA, но более слабая (или вообще отсутствующая) на рефлексы группы I (Jankowska 2010)
Модуляция рефлексов в ходе локомоции
И H-рефлексы, и GTO рефлексы можно аккуратно измерять в ходе локомоции. И соответственно показано, что они очень сильно модулируются в строгом соответствии с циклом шага.
H-рефлекс икроножной мышцы отсуствует (подавлен) при проносе ноги (swing phase), начинает усиливаться при контакте пятки с опорой, и достигает максимума в фазе опоры (stance phase).
GTO рефлексы вместо классического «расслабляющего» эффекта в фазе опоры поддерживают возбуждение в мышцах экстензорах, причем не только «своей» мышцы, но и например икроножная усиливает возбуждение экстензоров бедра.
H-рефлекс, как описано выше — просто удобный способ измерить работу «основного» моносинаптического Ia рефлекса натяжения. Считается, что такая его модуляция позволяет стопе отработать резкие изменения опоры — рефлекс этот по функции своей стабилизирующий.
В противовес этому в поддержании равновесия Ia рефлекс модулируется почти до полного его гашения, и в позном контроле короткий рефлекс не участвует.
Механихзм модуляции H-релфекса мозжечком
Как ни удивительно, внятного ответа на вопрос о том, какими супраспинальными структурами производится модуляция спинальных рефлексов, до сих пор не было.
Интересной поэтому является очень свежая серия работ Matsugi Akiyoshi из Shijonawate Gakuen University, Осака, в которых он показал модуляцию H-рефлекса путем транскраниальной магнитной стимуляции мозжечка. Модуляция H-рефлекса икроножных мышц имела пик через 110мс после импульса, и было показано, что она осуществляется через интернейроны пресинаптического, но не реципрокного торможения. Этот профиль спинальных связей характерен для вестибулоспинальных и ретикулоспинальных трактов. Matsugi постулирует именно такой путь, хотя и несколько косвенными аргументами.
Sakihara Kotoe, также из Осаки, попробовал также делать оптокинетическую стимуляцию людям, во время таких TMS опытов с людьми, показав, что на фоне стимуляции вестибулярных ядер посредством дополнительной зрительной системы (отмечу, что описание оптокинетических стимулов вызывает вопросы, она не очищена от потенциальных других путей), латенция моторных реакций мозжечка сокращается, в то время как латенция кортикальных реакций остается неизменной.
Если верить этим японским данным, вскрывается сеть управления рефлексами:
Кора мозжечка — (глубокие ядра мозжечка) — вестибулярные ядра и ретикулярная формация — вестибулоспинальные и ретикулоспинальные тракты — спинальные, сегментные интернейроны — пресинаптическое торможение на мотонейронах
Однако я пока не могу привести лучших теорий модуляции спинальных рефлексов со стороны мозжечка. Транскортикальный путь представляется еще более кривой теорией.
Group II афференты. Снова веретена
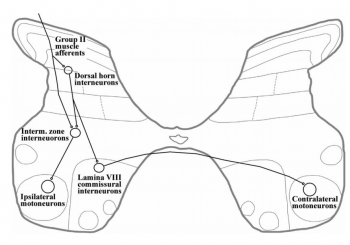
Несмотря на то, что афференты группы II от мышечных веретен не менее многочисленны, чем Ia, их роль часто просто игнорируется. Это связано с тем, что Ia реакции изучать легче — у них меньше порог, и в силу этого практически невозможно стимулировать только II группу, не затронув Ia. Почти всегда, когда говорят о веретенах, экспериментальные данные содержат преимущественно Ia эффекты.
Хотя описывают и моносинаптические связи, основной вид рефлекторной дуги II группы — через интернейрон. Внятной классификации интернейронов II группы нет, более того, это неоднородная группа. Однако во многих работах (включая Circ.Spinal.Cord) среди них выделяют проприоспинальные нейроны — то есть нейроны с обширными связями за пределами своего сегмнента.
Позная роль рефлексов II группы
Есть свидетельства того, что |Medium Latency (ML) ответ на растяжение мышц опосредован именно рефлексом II группы афферентов:
- он модулируется (усиливается, если мышца играет позную роль,и подавляется, если нет), в противовес SL моносинаптическим реакциям
- При болезни Шарко—Мари—Тута тип 1A преимушественно поражаются Ia афференты, и вместе с ними SL моносинаптические рефлексы, сохраняя ML рефлексы (хотя они и замедляются). Больные с таким поражением не испытывают проблем с балансом.
Circ.Spinal.Cord.p271-279